Introduction
Neural correlates of language and cognition in diverse populations, including bilinguals, have been of increasing interest and expanding to pediatric populations.1–3 Overall, behavioural evidence suggests that typically developing bilinguals achieve crucial linguistic and cognitive developmental milestones at a pace similar to that of monolingual children; however, a bilingual learning environment may also yield different trajectories.4–11 Whether potential differences between monolingual and bilingual development also exist at the neural level is unclear. Research has established various brain regions and networks relevant to adult bilingual language processing and cognition,12–14 however it is unclear when these bilingual activation patterns begin to emerge. It has been also proposed that it is critical to understand the impact of a richer – bilingual – language input on brain development and aging,15 neuroplasticity as well as,16 potentially, on cognitive reserve.17,18 Understanding the effects of a bilingual environment may be key to understanding the mechanisms influencing cognitive functioning across the lifespan and healthy brain aging. In the present meta-analysis, we explore the neural correlates of language and cognition during bilingual development.
Bilinguals vs. monolinguals
It has been a common research practice to contrast bilingual and monolingual outcomes, to gain a window into bilingual development. While these comparisons can be meaningful, it should be also considered that bilinguals represent a heterogenous group, in terms of their age of acquisition (AoA), proficiency, and current and past exposures to languages.19 Relevant here as well is that first (L1) and second (L2) language acquisition may be constrained to selected time windows or sensitive periods.20,21 For this reason, simultaneous bilingual children – those exposed to two or more languages since birth or prior to age three,22 though some classifications extend this range to beyond23,24 – versus sequential bilingual children – those exposed to another language after age three – might exhibit different neural developmental patterns. Here, bilingual is used inclusively to refer to both bilingual and multilingual language environments.
Language and cognition
In our analyses, we consider language and cognition in tandem to gain a comprehensive and ecologically valid understanding of the mechanisms underlying bilingual development. We operationalize cognition broadly to encompass higher-order mental processes, including but not limited to attention, memory, and executive functions (such as inhibition, working memory, and cognitive flexibility), as measured through both verbal and non-verbal cognitive tasks. Most theoretical schools accept that language is integral and a central part of cognition,25–27 given a deep interconnection between language abilities and broader cognitive processes.28,29
Regarding language, our scope includes tasks that focus on both sub-lexical and lexical (e.g., phonemic, syllable, word-level tasks) and supra-lexical (e.g., sentential, continuous speech) processes. This distinction captures the unit of linguistic processing, ranging from fine-grained acoustic-phonetic and lexical processing to higher-order syntactic, semantic, and discourse-level integration30–33 and may be particularly relevant for bilingual populations.34 It is also important to note that in most experimental paradigms reviewed here, there is a considerable overlap in measurements of language and cognition – particularly in task-based assessments of language (e.g., semantic/phonological interference task) and language-based assessment of cognition (e.g., phonological working memory task).
Language and cognition in the monolingual brain
Previous monolingual-focused, developmental neuroimaging studies have identified brain regions that we now commonly refer to as “classic language areas”, including the superior temporal gyrus (STG) and the left inferior frontal gyrus (IFG).35–37 Also, with increasing age, the language network develops greater left lateralization,38,39 (cf. recent findings suggesting adult-like lateralization by age 440) with increased activation of semantic processing regions (frontotemporal areas, e.g., classic language areas of Brodmann areas 45, 47 – IFG, fusiform gyrus, superior and middle temporal gyri – STG, MTG41 and sensory/motor regions (e.g., left premotor, bilateral primary motor, left somatosensory cortex37 and a decrease in recruitment of the default mode network (e.g., posterior cingulate cortex and precuneus) during cognitive control processes with age.37
Language and cognition: Monolinguals vs. bilinguals
In developing bilinguals, brain structure differences have been observed in comparisons with age-matched monolingual peers, albeit based on limited evidence.42 Across various adult and pediatric age groups, voxel-based morphometry studies revealed elevated levels of grey matter in bilinguals compared to monolinguals, in a solitary cluster located in the right culmen. Notably, the findings were largely influenced by the sole pediatric study included in the analysis, and when this study was removed, no significant differences were found in grey matter volumes between bilinguals and monolinguals.42,43
However, to date, no meta-analyses focusing on developing populations have compared the functional activations of task-based language and cognition, or the neural responses associated with performing specific language and cognitive tasks, across bilingual and monolingual children. While there have been meta-analyses of comparing adult bilinguals’ L1 and L2 performance on language and cognitive tasks,12–14,44,45 here we focus on monolingual versus bilingual comparisons. When drawing from adult literature reviews of the neural correlates of bilingual processing, there is no consensus on whether specific areas are engaged in processes unique to the bilingual experience.46–49 Both increased and decreased activity can be seen for the same area within- (across studies) and between-reviews. Greater control-related functional brain activity across multiple identified areas of anterior cingulate cortex (ACC), prefrontal cortex (PFC), putamen, caudate nucleus has been claimed for both monolingual and bilingual adults; this is also seen in the putamen/caudate nucleus for adult language learning. EEG studies on the bilingual N2 component activity, related to control, also show discrepancies, indicating both greater N2 – possibly indicating heightened inhibition/monitoring – and smaller N2 – potentially reflecting more efficient general processing depending on the task and population.47 Similar to the discussion on functional and structural connectivity as highlighted by García-Pentón,49 the “hazy view” of bilingual neural correlates arises from variations in analysis methods across studies and a heterogeneity in recruited bilingual samples, resulting in a lack of generalizability across results.
fMRI and fNIRS
Neuroimaging investigations in developmental research have successfully utilized functional magnetic resonance imaging (fMRI) and functional near-infrared spectroscopy (fNIRS). Both techniques provide valuable insights into dynamic changes during task-based brain activity. While fMRI typically offers higher spatial resolution and deeper penetration into brain tissue compared to fNIRS, the latter generally provides better temporal resolution. fNIRS especially has gained popularity in language studies involving pediatric samples due to its accessibility, versatility, and compatibility for use with children.50 While an important body of literature exist in this area, studies using electroencephalography were not included in the present review due to the limited spatial resolution of this imaging technique and given our focus on the neuroanatomy of bilingual processing.
Research questions
-
What brain areas are activated when typically developing bilingual children engage in language and/or cognitive tasks?
-
How do these activated brain regions in bilingual children compare with those identified in monolingual children from the same studies?
Methods
Two-stage systematic search
Study protocol preregistration is available here (https://osf.io/8NKWZ/). We adhered to the Preferred Reporting Items for Systematic Reviews and Meta-Analyses guidelines (PRISMA).51 Following consultation with a health science research librarian, we conducted a systematic search across three electronic OVID databases, Medline, PsycINFO and Embase. Concepts and more detailed search terms adapted for each database are available in the Supplementary Material (S1). In addition, a supplementary manual search using the reference lists of pre-qualifying articles from the electronic database search and relevant articles identified through Google Scholar ‘Cited By’ section was conducted. The most recent search was finalized in May 2024.
Selection criteria
Studies were included if: (a) they were peer-reviewed, primary texts written in either English or French without a specific date range; (b) they included participants <18 years; (c) the participants were identified as bilinguals in the article, including either simultaneous bilinguals (who have been exposed to both oral languages from birth up to three years of age) or sequential bilinguals (exposed to each oral language after the age of three); (d) the studies reported activations for language and/or cognitive tasks for each language group; (e) studies did not evaluate literacy (no reading-related task as to not exclude children at earlier ages); (f) the studies used one or more of these specific types of neuroimaging – fMRI or fNIRS; (g) the studies reported Talairach or MNI coordinates for each activation. No limitations were placed on the specific characteristics of the bilinguals.
Article screening
Both database and manual search results were subject to article screening. After duplicates were removed, a two-step screening process was applied. First, a screening of titles and abstracts was conducted by a team of trained reviewers (three trained graduate research assistants and author), with each article reviewed by two reviewers. Disagreements were resolved by a third reviewer who had not previously assessed the article. Second, a full-text screening was conducted with a focus on appraising the study critically using the “Checklist for Quasi-Experimental Studies” from the Joanna Briggs Institute Appraisal Tools.52 Again, two reviewers were tasked with appraising each study for potential for bias in methodology and results. After the full text of each article was screened, disagreements were once again reviewed by the third reviewer to determine inclusion into the final count. Once the disagreements were sorted, studies that qualified were added to the final list of articles included in the knowledge synthesis.
Critical appraisal of studies
The Joanna Briggs Institute Appraisal Tools “Checklist for Quasi-Experimental Studies” was adapted for appraising studies for risk of bias.52 Critical appraisal questions are available in the Supplementary Material (S2). Queries from the original tool checklist were followed, and additional questions were also added, tailored to the needs of this study:
-
Measure of socioeconomic status (SES) as matched between groups: As SES is an important factor in the study of bilingualism, measuring and matching SES between groups would be ideal in the articles selected (i.e., recruitment from the same neighborhood, parental education, household income).
-
Assessment for bilingual language background: studies should assess bilingual language background, at least identifying two factors: age of acquisition and proficiency.
To evaluate the assessments quantitatively, we transformed the responses (Yes, Unclear, No, N/A) to numerical values (100, 50, 0, N/A) and calculated summary statistics (average and standard deviation). We set an average score of >60 as a cutoff to only retain studies of medium and high quality.
Data extraction and syntheses
The following data was extracted from included studies: study location, age, language background history, neuroimaging technique (fMRI, fNIRS), nature of the task performed (task), linguistic or cognitive level of processing (sub-lexical and lexical – phonemic, syllable, word; supra-lexical – sentential, continuous speech; cognitive – executive function, social cognition), name and location (x,y,z) of regions showing significant activity, and summary of main findings.
For the fNIRS studies, given the heterogeneity in task design, fNIRS systems, optode placement, data preprocessing, and reporting standards (see Supplementary Material S3), it was determined that a meta-analysis could not be conducted. While previous meta-analyses have incorporated fNIRS studies by accounting for differences in methodology, the level of variability in the present dataset – particularly in data acquisition and the definition of regions of interest –posed significant challenges for synthesizing effect sizes in a meaningful way (see the Limitations section for more details). As a result, a narrative synthesis was deemed the most appropriate approach to summarize the findings. Accordingly, we present a narrative synthesis that integrates findings from both fNIRS and fMRI studies, alongside a meta-analysis of the fMRI data.
For the fMRI studies, an exploratory fMRI analysis was conducted through activation likelihood estimation (ALE)53; using GingerALE software, version 3.0.2 (http://brainmap.org/ale/). 3D maps were generated via the collected locations (x,y,z) of regions. The likelihood of significant activation within a given voxel is represented by ALE scores resulting in activation maps and compared to a null distribution54,55 and a random effects model accounts for clustering between experiments to limit the effect of a single experiment. All contrasts and foci from fMRI articles were manually extracted and input into the BrainMap’s Scribe database. Coordinates reported in Talairach were converted to Montreal Neurological Institute (MNI) standard space in GingerALE. Given the exploratory nature of this analysis, we used an uncorrected threshold of p<0.001, with a minimum volume of 150 mm3, comparable to previous ALE meta-analyses investigating bilingual subjects.12,13,44,56 Voxels surviving this threshold were interpreted as showing above-chance convergence between experiments reflecting the language- and cognitive-associated activations in children. Thresholded ALE maps were overlaid onto the Talairach coordinate brain template,57 using image viewing software Mango, version 4.1 (http://rii.uthscsa.edu/mango/).
The following analyses were performed: (i) within-group (bilingual) activation patterns during language and/or cognitive tasks; (ii) between-group (bilingual ≠ monolingual) activation patterns during language and/or cognitive tasks.
Results
First, we present a narrative synthesis of the results from both fNIRS and fMRI studies. We then present an exploratory ALE meta-analysis of fMRI studies, given the limited number of available studies (n=6).
Systematic search
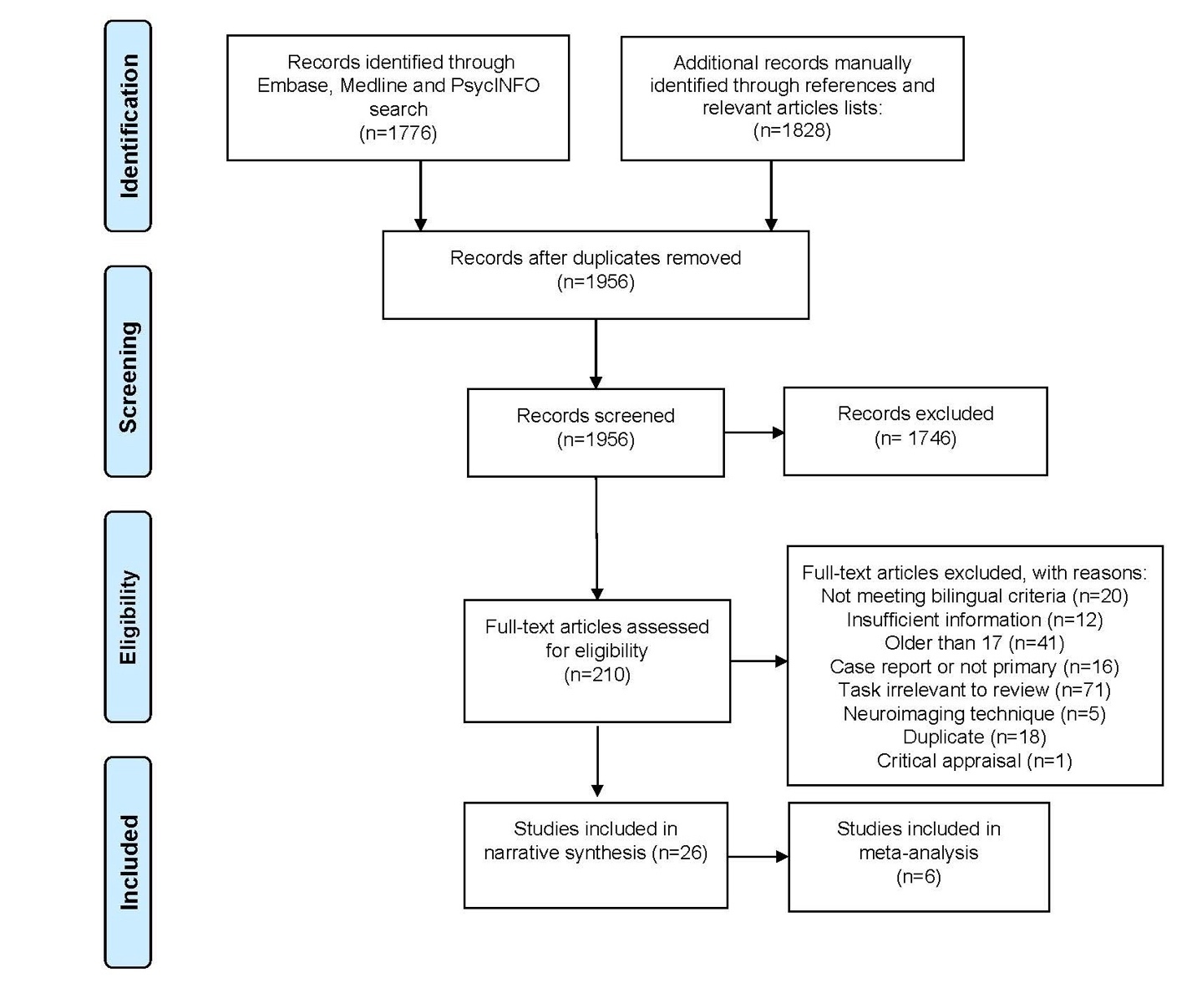
Twenty-six studies met inclusion criteria, of which 6 included fMRI data; those were analysed in an exploratory meta-analysis. Figure 1 provides a summary of the search, number of study exclusions to final inclusion following the PRISMA flow diagram.51 Articles were excluded for the following reasons: did not include bilinguals according to the defined criteria; missed sufficient information (e.g., activation reported separately by group, coordinates); did not recruit children (participants older than 17 years); not a primary study or a case report; reported on tasks that were beyond the scope of the current review (e.g., reading tasks); did not use fNIRS/fMRI neuroimaging and additional duplicate records were missed in the original duplicate screening. After evaluating the quality of the studies, one more study was excluded from synthesis. Details on study characteristics for fNIRS and fMRI studies are available in Table 1 and Table 2 respectively.
Critical appraisal
Critical appraisal assessments were conducted for all studies that met inclusion criteria (S2). We excluded one study58 from our synthesis as it fell below the established threshold (score >60; medium and high quality). Although other studies also lacked information on either SES or bilingual language background, the decision to exclude this study was based on its inadequate reporting of both elements, with no details provided on SES or bilingual background, causing it to fail to meet our threshold.
_flowchart_for_.jpeg)
Participants
Across the 26 studies, a total of approximately N=1,460 participants were included. All studies recruited bilingual participants (n=928). Twenty studies included at least one comparator group, most often age-matched monolinguals (19/20; n=532) or more rarely, another bilingual group (2/20; e.g., different age groups, different language pairing). Some overlap across samples is possible, particularly in longitudinal or lab-based studies, and therefore this number should be interpreted as an approximate upper bound of unique participants.
Age at test
Most participants were recruited at two developmental stages, around the start of kindergarten (3-6 years old) and during early elementary school (7-12 years old). However, the age range of the participant sample was wide, ranging from 4 months to 16 years, with a mean age of 7.06 years (standard deviation = 3.72) for bilinguals and 6.95 years for monolinguals (standard deviation = 4.22). Three fNIRS studies and one fMRI study recruited bilingual participants of multiple ages for the purpose of conducting analyses using age as a predictor variable. The fMRI study included two distinct age groups: 6-8 and 9-10-year-olds.59 The three fNIRS studies involved infants at 6-month-olds and 10-month-olds,60 and 4-6-month-old and 10-12-month-old infants61; the other study recruited an older school age group of 6–8-year-olds and 9-11-year-olds.62
Bilingual type
Given our definition of exposure to two languages before three years of age, simultaneous bilinguals were most often reported (n=10), whereas sequential bilinguals were reported less often (n=3; Table 1-2). Only one study compared performance across bilingual type, recruiting both simultaneous and sequential bilingual groups.63 Studies also recruited a mixed group of simultaneous and sequential bilinguals, often labeled “early bilinguals” (n=10; Table 1-2), given the fact that authors may have used different age cutoffs for designation of bilingual type (e.g., age four or five). Unfortunately, a subset of studies did not provide enough information to determine bilingual type (n=4); these children were often described as learners of English as a foreign language.
Languages of tasks and languages used by bilinguals
Many studies used an English-language task, as the most often spoken language across studies was English. However, a few research groups used French64,65 and German.66,67 Several studies conducted the task in both bilinguals’ languages (8/26). Bilinguals were almost always heritage language speakers (e.g., language learned from home environment), learning an additional societal language, or the language commonly used as a majority language in the society of the speaker (e.g., English and French in Canada, German in Germany). While the term ‘heritage speaker’ can imply broader patterns of language shift or attrition (see Benmamoun et al.68), the studies reviewed here focus on individuals still in the process of language acquisition and do not describe such pattern. The most often reported bilingual heritage language was Spanish, followed by Mandarin Chinese, and Japanese. Studies most often recruited homogenous groups of bilinguals, where all children had in common the same language pairings (e.g., all speakers of English and Spanish), as seen in 23/26 studies (Table 1-2). One study recruited two homogenous groups of different language pairings.69 The remaining three studies recruited heterogenous groups of bilinguals, where all participants shared one language (societal language, language the task was conducted in) but differed in their other language, often a home/heritage language.
Study location
Half of the included studies were conducted in North America (United States and Canada). Additional sites include European (Germany, France and United Kingdom) and East Asian nations (Taiwan, China and Japan).
Methodology
Neuroimaging technique and appropriateness for meta-analyses
fNIRS studies (n=20) largely outnumbered fMRI studies (n=6), given the enhanced applicability of fNIRS in pediatric samples and its lower cost compared to MRI. Methods for reporting fNIRS signal acquisition (e.g., fNIRS instrument, source-detector distances, regions of interest and laterality) were heterogenous across studies; fNIRS signal acquisition details are available in Supplementary Material (S3). Given a lack of openly available channel-based data, we found the fNIRS studies were not suitable for a meta-analytic approach in the current study.70
As for fMRI, all 6 studies reported BOLD signal imaging obtained through 3T MR scanners, with coordinates in MNI space. fMRI protocols varied in terms of data acquisition parameters and pre- and post-processing (software, preprocessing steps). Stimuli were presented through insert headphones (n=3) or not otherwise specified. Findings are explored using an ALE meta-analysis approach in the following section.
Experimental designs and tasks
Auditory language-related tasks were most employed, often using sub-lexical and lexical (e.g., phonemic, syllabic, word-level processing; n=10) and supra-lexical (sentence-level, continuous speech processing; n=10) listening tasks and morphological awareness tasks (Table 1-2). Six out of the twelve cognitive tasks were language-based cognitive tasks, evaluating executive functions (working memory, monitoring, switching) through tasks including phonological working memory (n-back), lexical competition, bilingual comprehension switch and property term learning forced choice. The other half were cognitive-only paradigms, typically evaluating executive functions (attention, switching) using tasks such as the Attentional Network Task and Dimensional Change Card Sort Task. Mode of task stimulation skewed towards active tasks that required participants to think about, process information and/or provide a response (e.g., button press), as opposed to passive (n=5), minimally demanding, listening-only tasks.
Group comparison: Bilingual (simultaneous, sequential/mixed)
The activation patterns presented below are derived from a qualitative synthesis of fNIRS and fMRI findings across simultaneous bilinguals, as well as other bilingual types (e.g., sequential, mixed). These patterns are reported separately for sub-lexical and lexical language, supra-lexical language, and cognitive tasks. Although the observations reflect trends we have identified across studies, they should be interpreted with caution due to considerable methodological variability and the limited number of available studies (see Limitations section for further discussion).
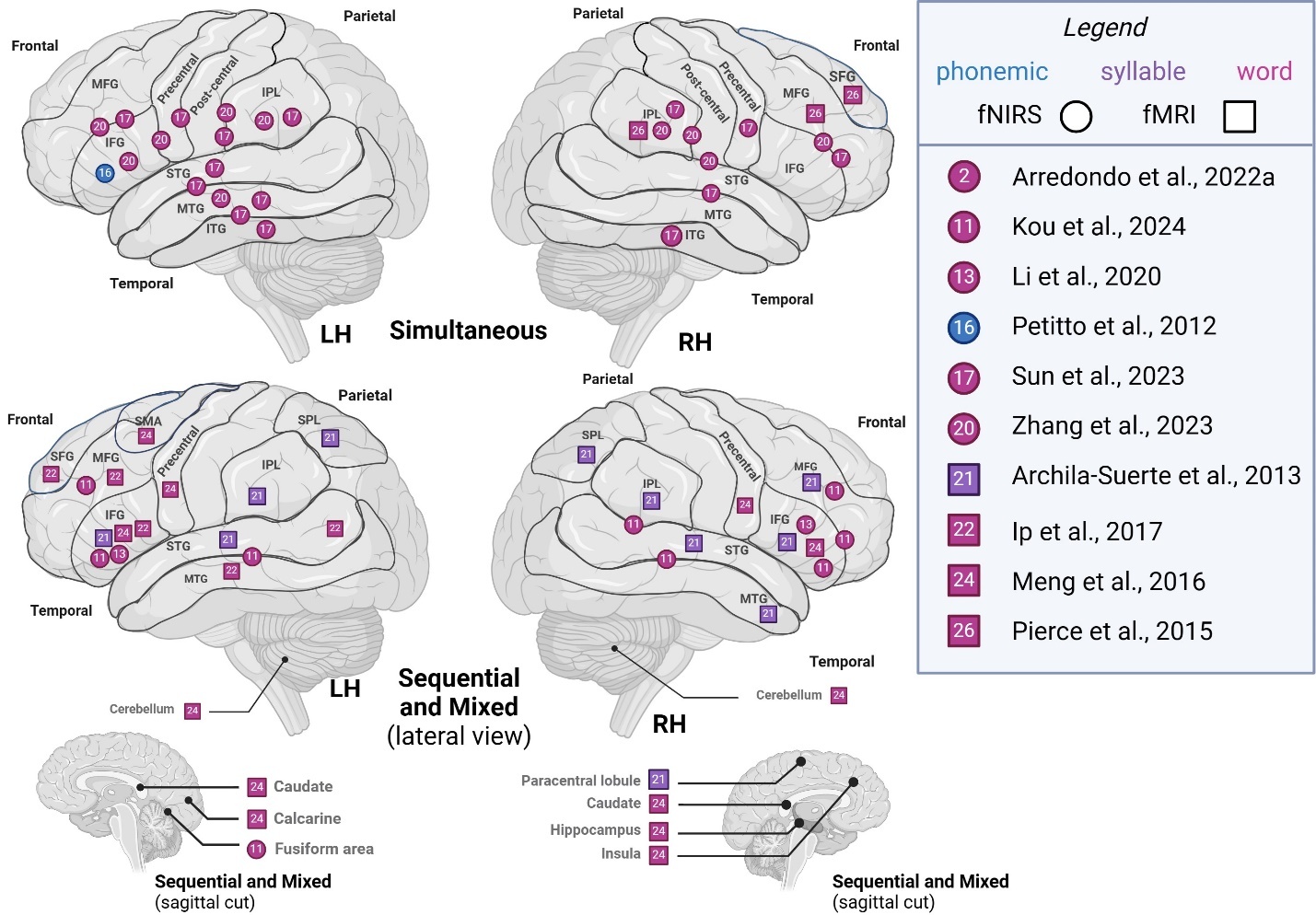
Sub-lexical and lexical (phonemic, syllable, word)
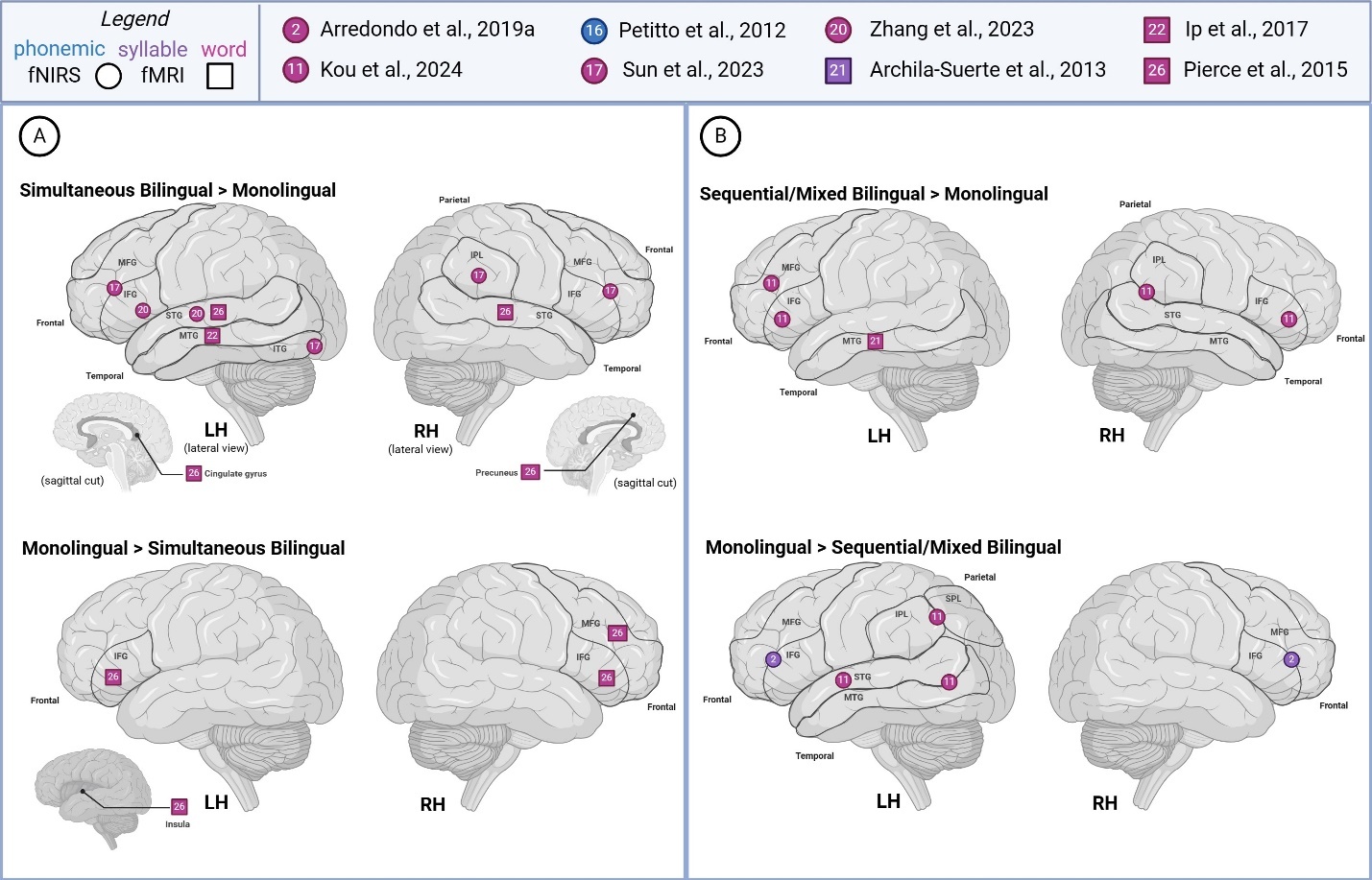
Comparing studies recruiting simultaneous bilinguals to other bilingual types (sequential, mixed), we represented segmental level activation patterns across hemispheres in Figure 2. In phonemic- and word-level studies, left hemisphere (LH) activity in simultaneous bilinguals was generally localized to frontoparietal and temporal areas. In contrast, LH activity in other types of bilinguals appeared more diffuse, with additional activations reported in superior frontoparietal regions, as well as in cerebellar, medial surface, and subcortical areas across phonemic-, syllable-, and word-level studies. Right hemisphere (RH) activity in simultaneous bilinguals was similarly localized to frontoparietal and temporal areas. In contrast, other types of bilinguals showed additional activations in cerebellar, medial, and basal surface regions, as well as subcortical structures such as the paracentral lobule, fusiform gyrus, caudate, hippocampus, and insula. Overall, simultaneous bilinguals tended to report a more constrained pattern of activation on frontal and parietal lobes’ superolateral surface, compared to studies that recruit sequential or mixed groups which often report more diffuse or additional activations.
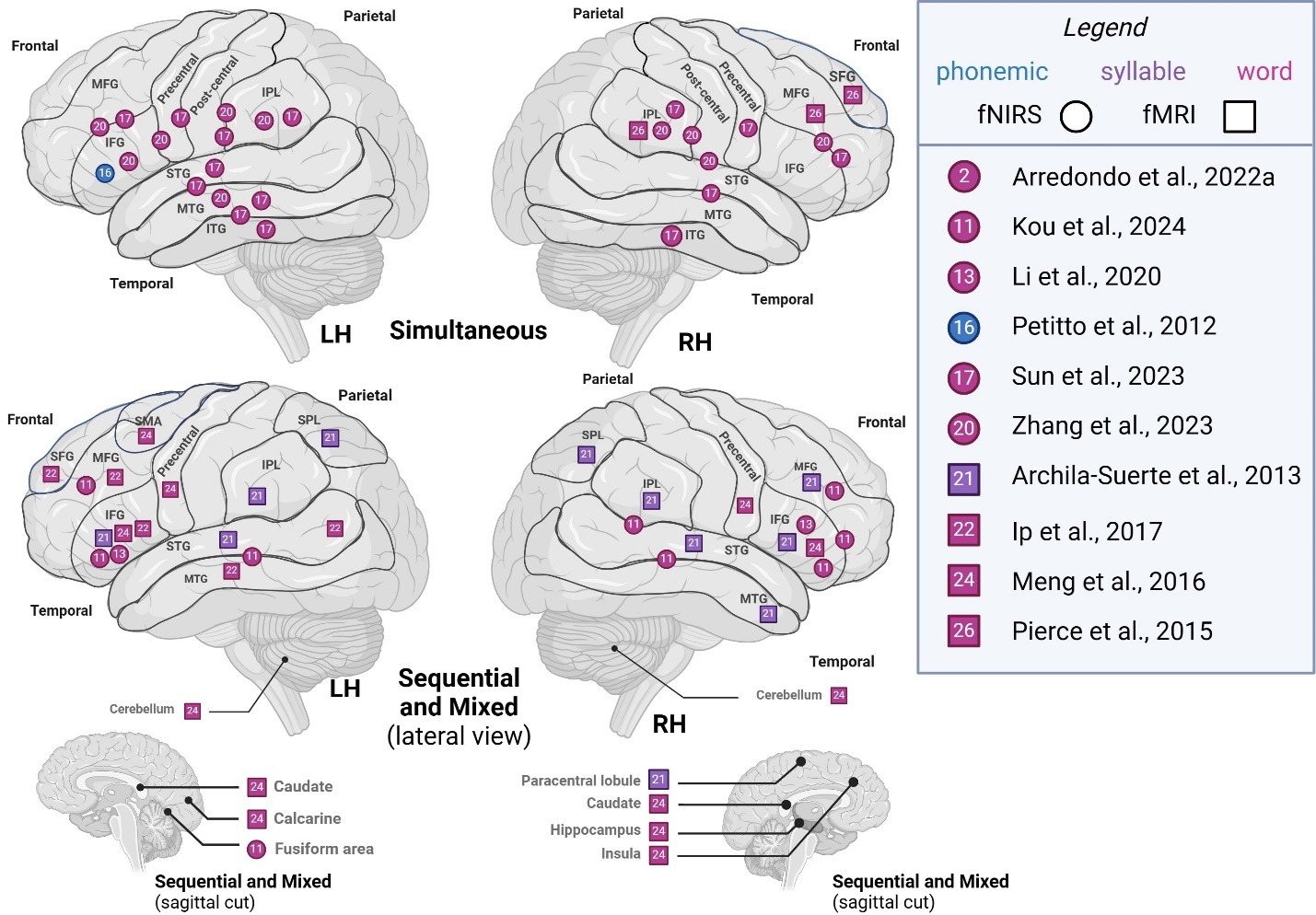
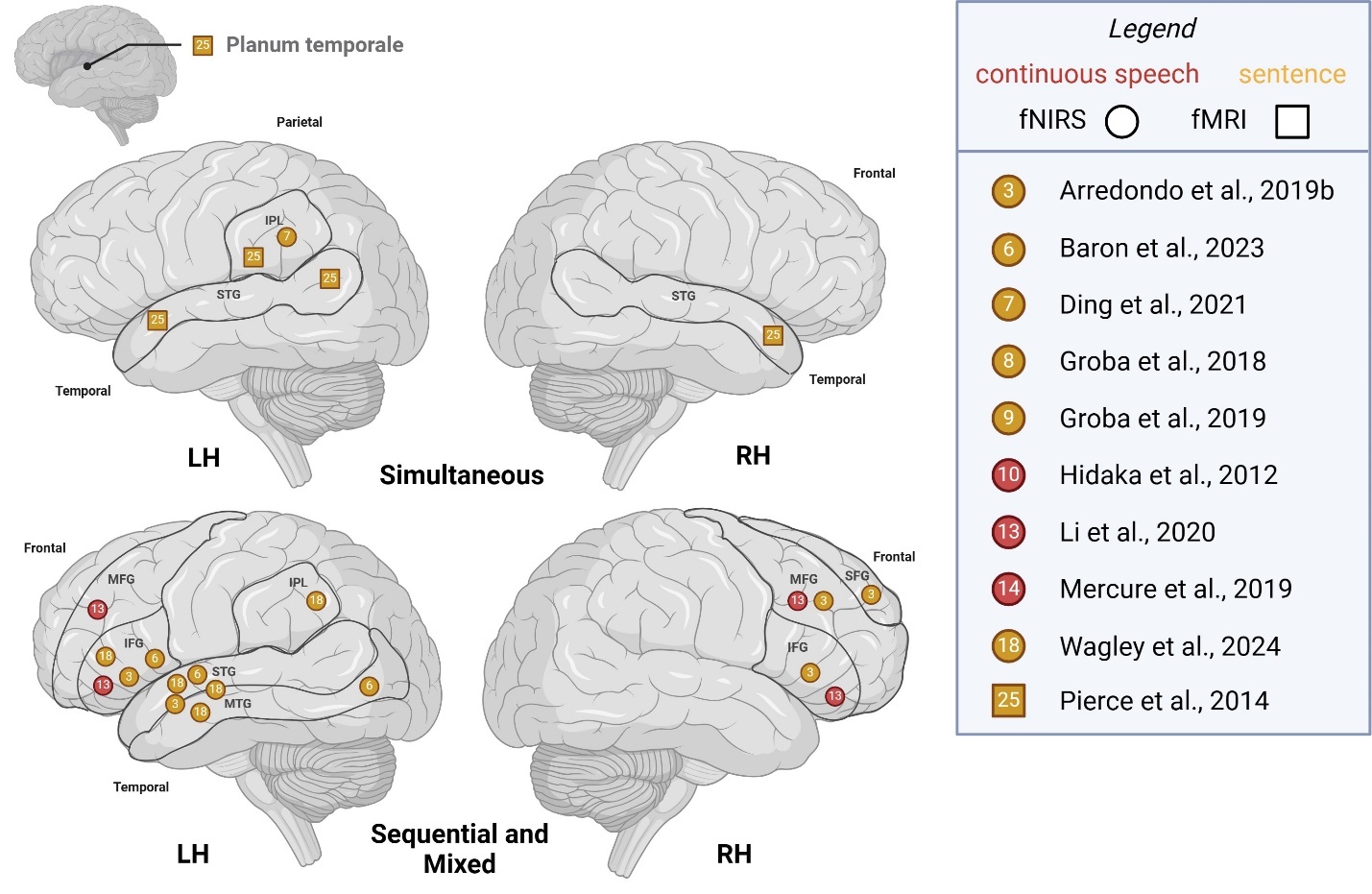
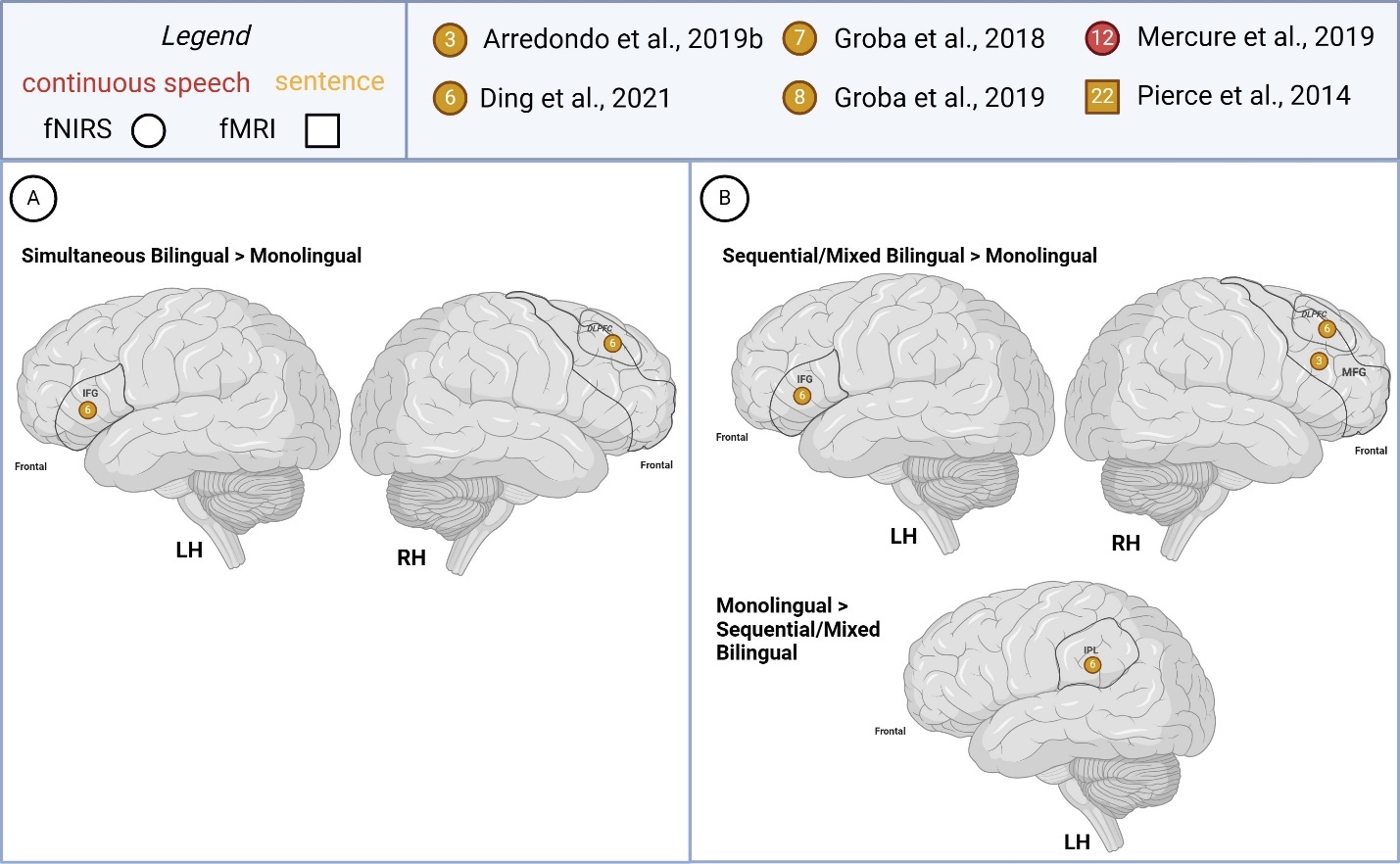
Supra-lexical (sentence, continuous speech)
To compare studies recruiting simultaneous bilinguals with those including other bilingual types (sequential, mixed), we illustrated supra-lexical-level activation patterns across hemispheres in Figure 3. For simultaneous bilinguals, LH activity tended to be observed in the IPL, posterior temporal cortex and planum temporale for sentence-level tasks, whereas reported LH activity for other bilinguals additionally included frontal areas of MFG, IFG for both supra-lexical level tasks. RH activity for simultaneous bilinguals was noted in the DLPFC, STG/MTG and posterior temporal cortex, whereas RH activity for other bilinguals included the SFG, MFG and IFG.
_areas_of_activation_a.jpeg)
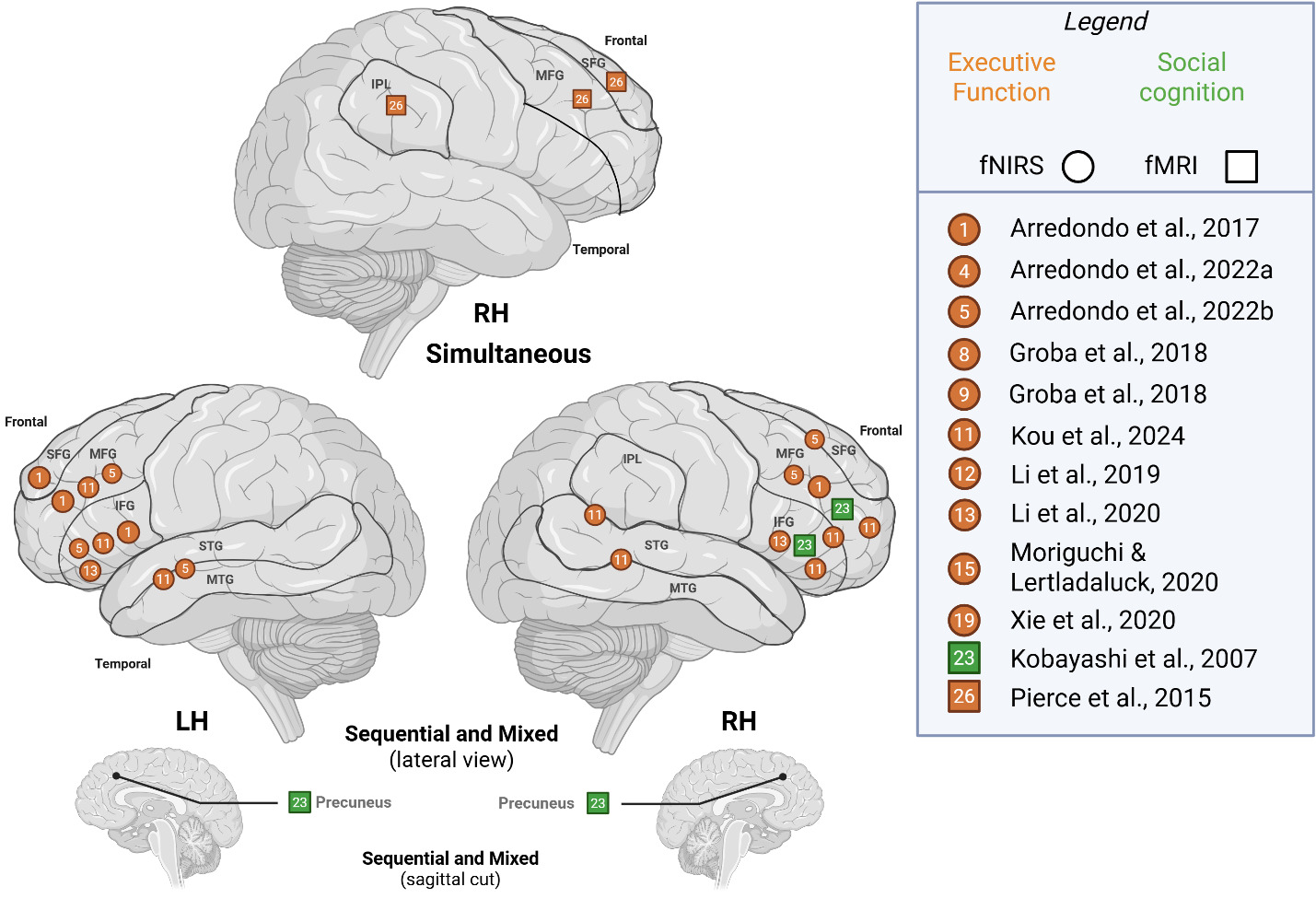
Cognition (executive function, social cognition)
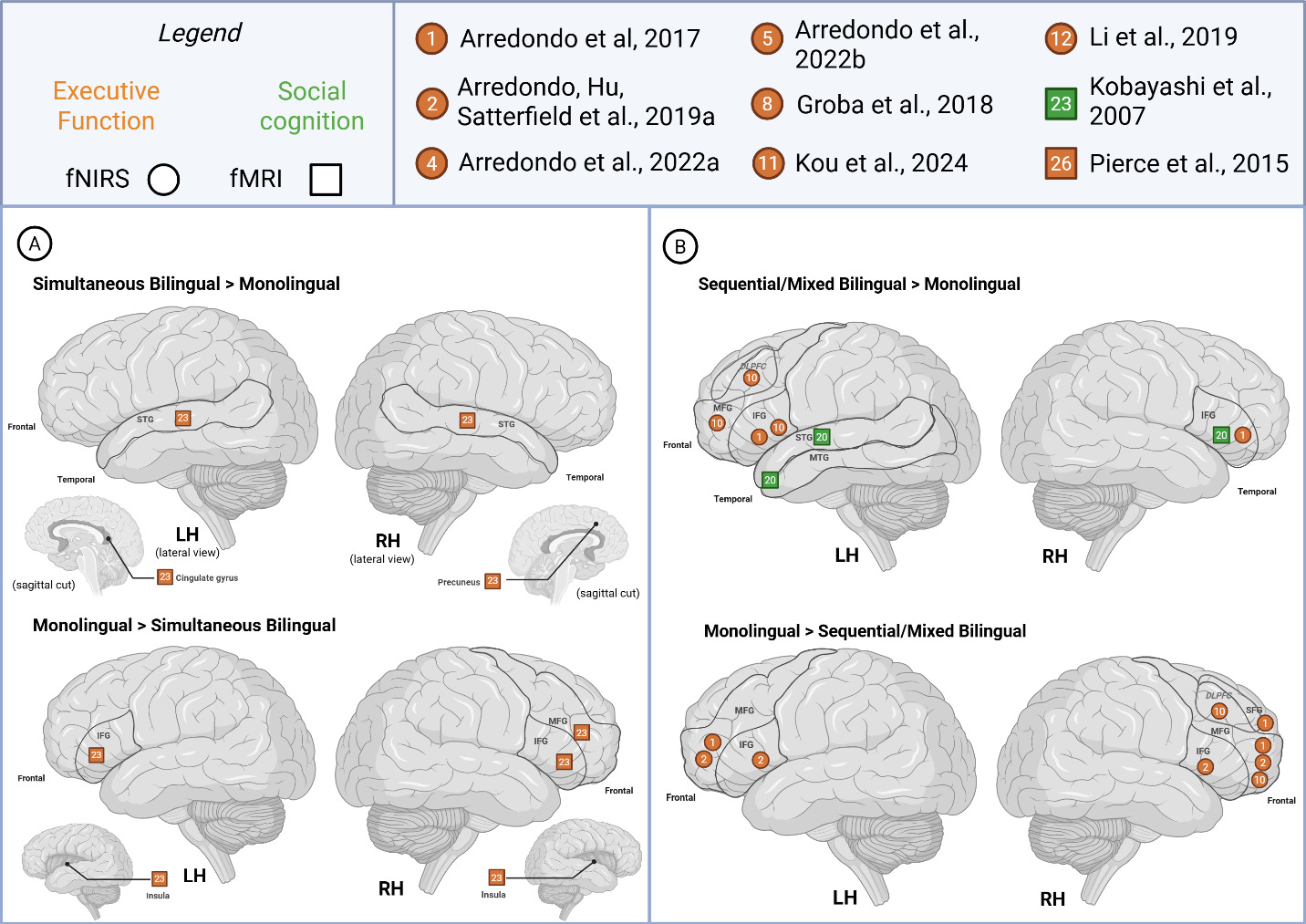
To compare studies recruiting simultaneous bilinguals with those including other bilingual types (sequential, mixed), we illustrated cognitive-level activation patterns across hemispheres in Figure 4. For simultaneous bilinguals, RH activity for was noted in IPL, SFG and MFG, whereas other bilinguals activated frontal areas – SFG, MFG, IFG – with additional reports of activity in the STG/MTG, IPL/STG and subcortically in the precuneus. LH activity was only reported by studies that recruited other bilingual types and largely activated frontal areas and STG/MTG; thus, we are not able to make comparisons on bilingual type.
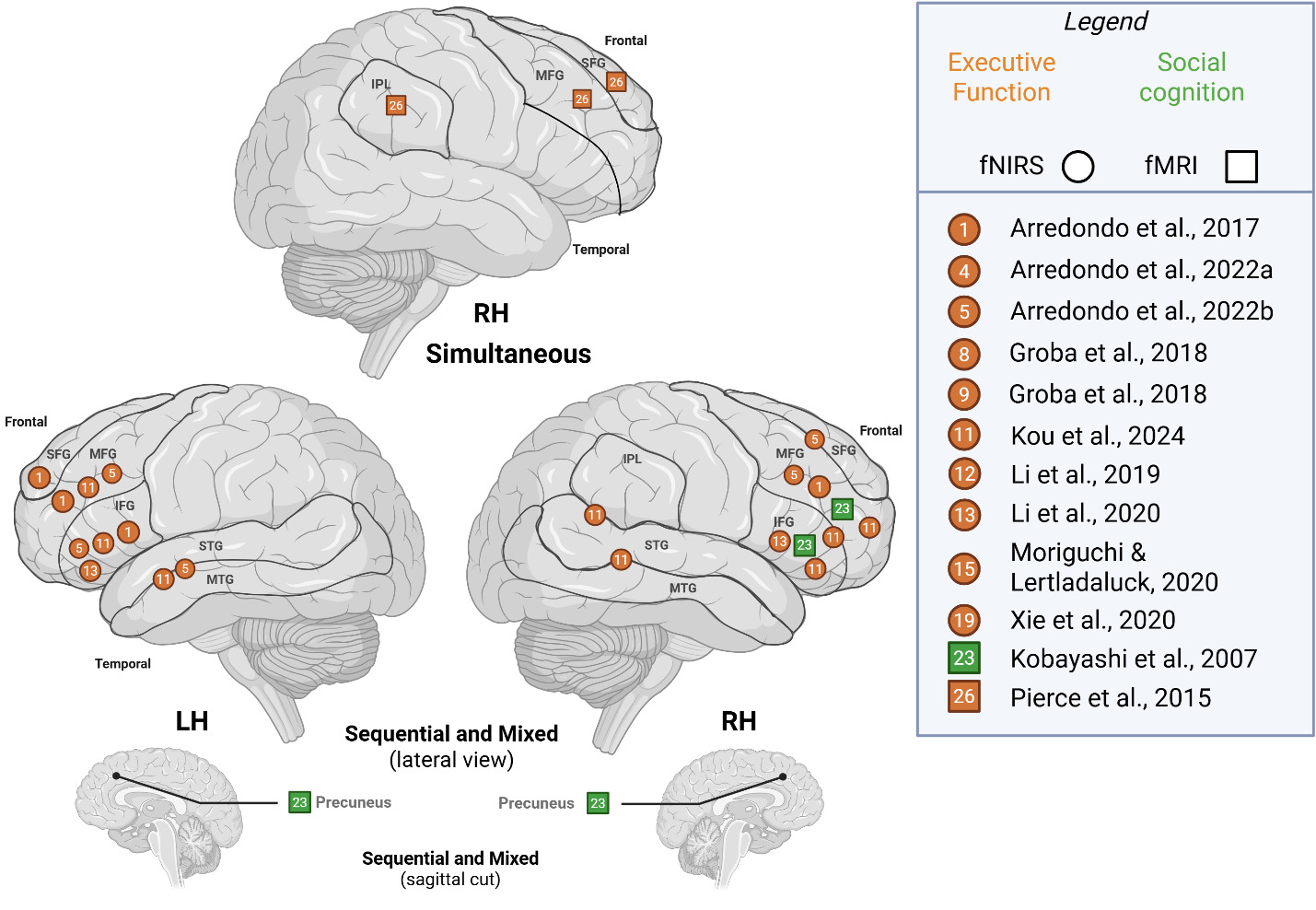
Group comparison: Bilingual (simultaneous, sequential/mixed) vs. monolingual
Nineteen studies evaluated monolingual and bilingual (simultaneous, sequential/mixed) children on both language and cognitive tasks. The results are presented separately for sub-lexical and lexical, supra-lexical, and cognitive tasks, with comparisons made between each bilingual group (simultaneous, sequential and mixed) and monolingual counterparts. As noted earlier, patterns are presented for descriptive purposes only and should be interpreted with caution.
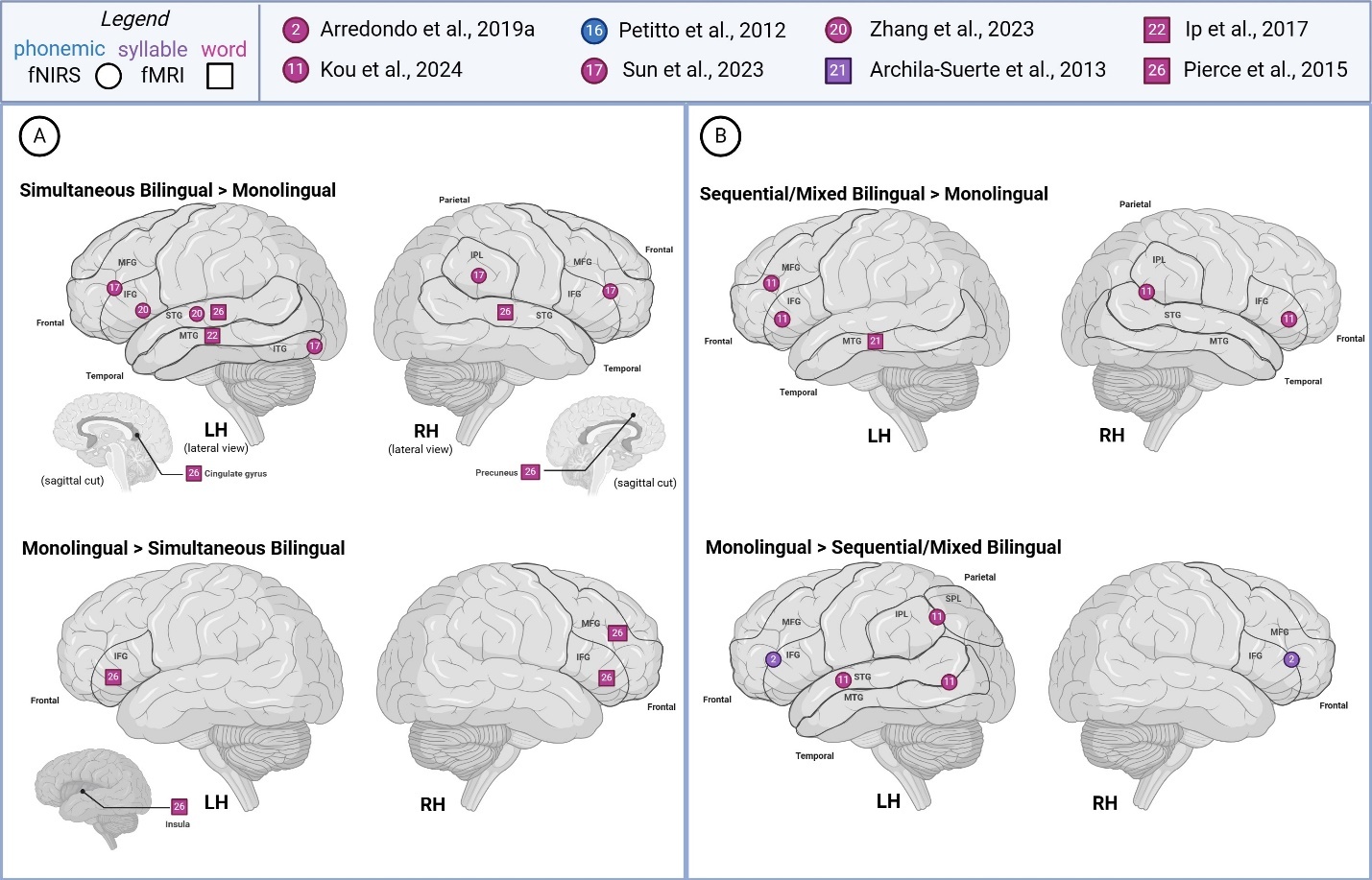
Sub-lexical and lexical (phonemic, syllable, word)
Figure 5 summarizes reported findings from sub-lexical and lexical language studies comparing bilingual and monolingual children. Sub-lexical and lexical language studies generally reported greater LH activity across all groups, with a tendency for higher reported activity in simultaneous bilinguals (>monolingual) during word-level task processing. For the most part, syllabic and word-level studies reported increased activation in distinct brain regions for simultaneous and sequential/mixed bilinguals, in comparison to monolinguals – though both bilingual groups demonstrated greater activity in the left MTG. Simultaneous bilinguals were noted to show higher activation in bilateral STG, left MTG, left cingulate gyrus, and right precuneus. Sequential/mixed bilinguals were reported to exhibit increased activation in the left MFG, bilateral IFG, left MFG, and right IPL/STG. Monolinguals were observed with higher activation in the bilateral IFG (> simultaneous bilingual), area bordering MFG/IFG, right MFG, IPL/SPL (> sequential/mixed bilingual) and temporo-occipital area and left insula (> simultaneous bilingual). Interestingly, all comparison groups reported increased activity in the left IFG, while different groups also reported higher activity in the MFG/IFG border area (monolingual > sequential/mixed bilingual; simultaneous bilingual > monolingual).
For the phonemic task,61 studied groups varied in their activation patterns, sometimes showing starkly opposite patterns for the same area. For instance, similar levels of LH IFG activity for younger monolingual and simultaneous bilinguals were reported, whereas older groups of simultaneous bilinguals (>monolingual) reported greater activation of the LH IFG. Considering both age groups, monolinguals showed greater LH activation of IFG than bilinguals.
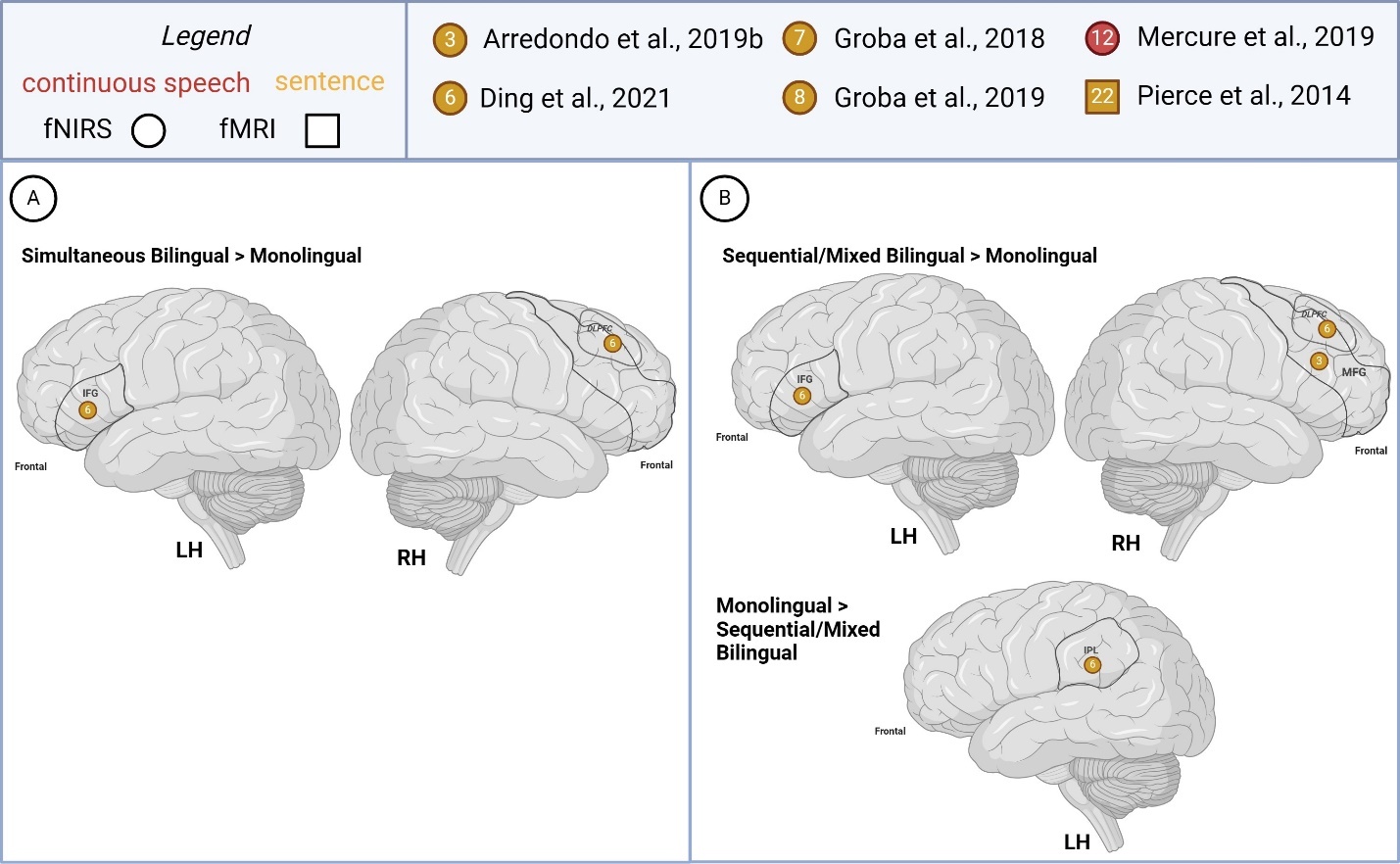
Supra-lexical (sentence, continuous speech)
Few studies (n=3) compared bilingual and monolingual children’s supra-lexical task-based activations, all pertaining to sentence-level processing (Figure 6). Findings between simultaneous and sequential bilinguals (>monolingual) were similar as one study recruited both groups Ding et al.63 in which greater LH IFG and RH DLPFC was reported in both bilingual groups (> monolingual). The left anterior temporal cortex and right MFG activity was reported as greater in sequential bilinguals (> monolinguals).
One study reported opposite group difference patterns for the different study conditions examined.71 For instance, bilateral IFG activity was reported as greater in simultaneous bilinguals (>monolinguals) overall but also in monolinguals (> simultaneous bilinguals) when considering findings from the higher attentional demand/difficult sentence type condition only.
Greater RH IPL, temporoparietal activation was reported in monolinguals (> sequential/mixed bilingual). Right frontal activation was also reported as higher in monolinguals (> simultaneous bilingual).
Cognition
Cognition studies reported greater LH activity primarily in sequential/mixed bilinguals (>monolinguals) for executive function tasks across the frontal regions of MFG, (DL)PFC and bilaterally in the IFG (Figure 7). For social cognition tasks, the right IFG and left STG and temporal pole were reported as more active in bilinguals (>monolinguals). On the other hand, simultaneous bilinguals (>monolinguals) were noted to show greater activity bilaterally in the STG, left cingulate and right precuneus.
Studies of executive function reported higher activation for monolinguals (> both bilingual) in the bilateral frontal regions and insula. One study reported various hemispheric activation patterns, sometimes showing opposite patterns for the same area depending on the trial and contrasts studied.60 For instance, bilateral IFG activity was reported as greater in bilinguals (conflict > control + rest trial) and monolinguals (control > rest trial), depending on reported contrasts. Likewise, LH SFG activity was reported as greater in bilinguals (conflict > control + rest trial) and monolinguals (conflict > control; control > rest trials) in this same study.
The social cognition study reported higher activation for monolinguals (> sequential/mixed bilingual) in right temporoparietal junction.72 Other cognition studies also reported greater monolingual (> simultaneous bilingual) activity in the left PFC and temporoparietal region and right frontal. Increased activity has been reported in the bilateral IFG and left MFG among both monolingual (>sequential/mixed bilingual) and sequential/mixed bilingual (>monolingual) groups.
_simult.png)
Exploratory ALE meta-analyses
fMRI studies and contrast analyses included in the meta-analysis are seen in Table 3. Six studies used fMRI, comprising a total of N=127 participants (Bilingual, n=96; Monolingual n=31). The paucity of the studies did not allow a specific analysis on bilingual type. Three sets of contrasts were analysed:
-
Set 1: Bilingual contrasts - within-group (bilingual) activation patterns;
-
Set 2A: Bilingual>Monolingual contrasts - unidirectional between-group (bilingual and monolingual) activation patterns;
-
Set 2B: Bilingual>Monolingual and Monolingual>Bilingual contrasts, henceforth Bilingual ≠ Monolingual - bidirectional between-group (bilingual and monolingual) activation patterns.
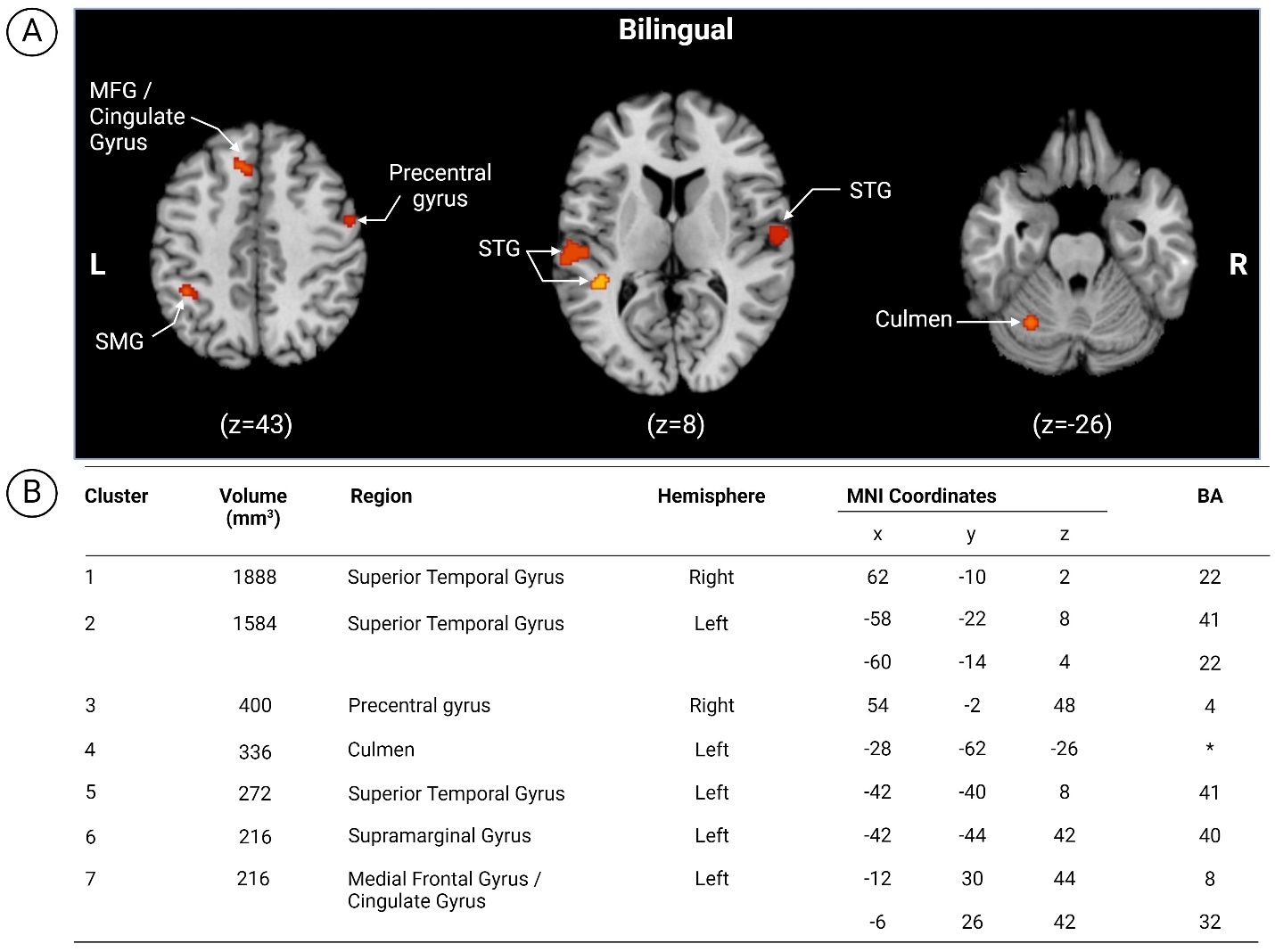
The first set of analyses targeted within-group (bilingual) activation patterns during language and/or cognitive tasks. Multiple fMRI contrasts reported within a single article constitute multiple independent experiments if they were obtained from different samples.59 Seven clusters emerged in the following regions of the left hemisphere (Figure 8): (i) STG; (ii) culmen; (iii) supramarginal gyrus (SMG) and (iv) MFG/cingulate gyrus and right hemisphere: (i) STG and (ii) precentral gyrus.
_thresholded_within-group_(bilingual)_ale_maps_with_mni_template_anatomical_underlay_sh.jpg)
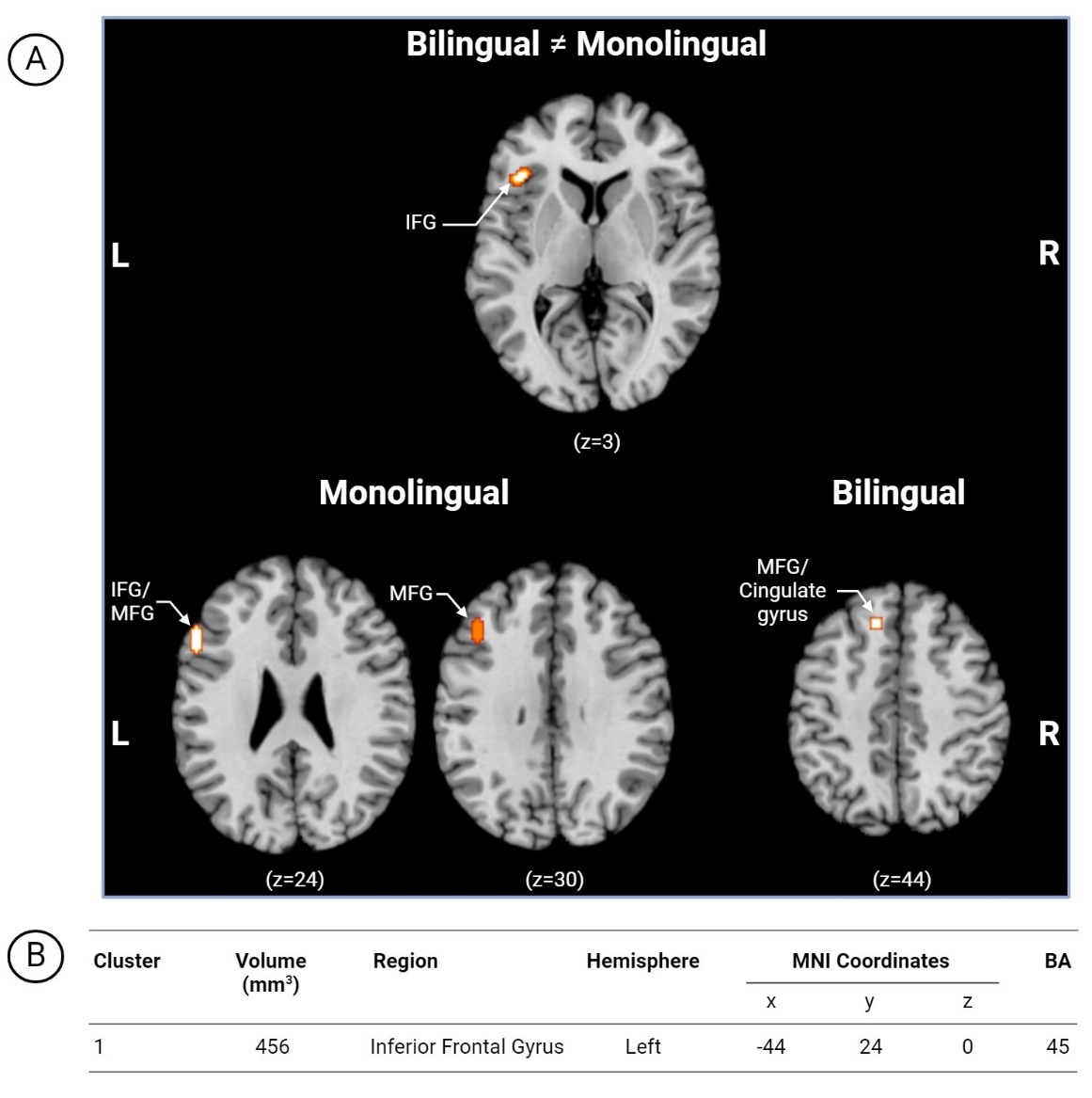
The second set of analyses focused on between-group (bilingual and monolingual) activation patterns during language and/or cognitive tasks. Bilingual > Monolingual contrast analyses found no significant clusters. Contrast analyses combining multidirectional between-group analyses (Bilingual ≠ Monolingual) were also explored. Multiple fMRI contrasts reported within a single article constitute multiple independent experiments if they are obtained from different samples, however samples were the same in these included papers. Contrasts were pooled into a single experiment to control for within-group effects.54,73 One cluster in the left hemisphere, inferior frontal gyrus (Figure 9) displayed activation.
_thresholded_between-group_(bilingual__monolingual)_ale_map_with_mni_template_anatomic.jpg)
Discussion
The present systematic review and exploratory meta-analyses was performed to describe the neural correlates of language and cognition in the developing bilingual brain. Following an extensive literature search, we identified 26 neuroimaging studies (20 fNIRS, 6 fMRI), comprising approximately N = 1,460 participants (bilingual: n = 928; monolingual: n = 532), with ages ranging from 4 months to 16 years. A subset of these studies also conducted group comparisons with age-matched monolingual counterparts (n=19).
Narrative synthesis
Studies comparing simultaneous bilinguals to other bilingual types show distinct patterns of brain activity across linguistic levels of processing. Our results show simultaneous bilinguals exhibit more localized LH activity in frontoparietal and temporal areas for phonemic and word-level tasks, while sequential/mixed bilinguals show more diffuse activation, including additional cerebellar, medial surface and subcortical activity. Supra-lexical and cognitive studies reveal similar trends, with simultaneous bilinguals showing specific patterns for LH and RH activations, compared to other bilingual types.
Our findings suggest that bilinguals rely on “classic language areas” previously defined by monolingual studies, such as the left IFG, from a young age, commencing as early as 4-6 months in the youngest bilinguals included and persisting throughout the lifespan. Across all linguistic levels of processing, both IFG and STG have been consistently highlighted as key areas for bilingual language and cognition, showing left dominant but bilateral activity. The IFG especially is important to both monolingual and bilingual processing as both groups claimed greater activity in this area depending on the study/level of processing examined compared to each other. This suggests that IFG activation in the context of language and cognition may reflect contributions from both core language processes and domain-general networks, such as executive functions, a point increasingly recognized in the literature (e.g., Fedorenko and Blank74). These findings highlight shared and unique mechanisms between bilingual and monolingual individuals, with the divergent findings regarding greater monolingual and bilingual processing in the IFG warranting investigation as more studies become available.
Language tasks also engaged frontoparietal control areas, such as DLPFC, IPL and other areas surrounding the intraparietal sulcus, traditionally associated to cognitive functions. It is possible this activation pattern is influenced by the study design as a greater proportion of active overt-response tasks were chosen over passive-listening tasks. These active tasks were varied but required bilinguals to process and monitor linguistic information and select between competitors – all of which would also depend on cognition as well. Active-response studies in monolingual children have also required greater memory and attentional resources, with greater recruitment of a wider network of additional areas and a larger effect size in left IFG compared to passive tasks.75,76 These results are also consistent with adult bilinguals’ language and task switching, with language-related activations across multiple areas, including the IPL, DLPFC, IFG – albeit all in the left hemisphere.14
Comparing language and cognition-related activation across monolingual and bilingual (simultaneous, sequential/mixed) representation, a pattern emerged across studies of sub-lexical and lexical task processing, whereby simultaneous bilinguals (> monolinguals) primarily reported greater frontal and temporal activity. Supra-lexical studies, while few, also corroborated this pattern. Cognitive tasks were associated with greater activity for sequential/mixed bilinguals (>monolinguals) on the other hand, also concentrated in the left frontal and temporal regions. Results show a closer alignment between sequential/mixed bilingual and monolingual groups especially evident in cognitive tasks, as both groups claimed higher activity in the same left frontal regions compared to each other. This pattern across bilingual types sheds some light on the effects of experience-dependent functional effects, relating to early vs. later language exposure. This is concordant with theories like interactive specialization,77 which posits that initially brain regions have broad responsiveness to various stimuli and tasks and then over time, through competitive and cooperative interactions, these regions become more specialized and interconnected.
Meta-analysis
Seven clusters emerged from the within-group analysis of bilingual contrasts highlighting regions of the left hemisphere including (i) STG; (ii) culmen; (iii) SMG and (iv) MFG/cingulate gyrus as well as the right hemisphere: (i) STG and (ii) precentral gyrus. Compared to other adult reviews focusing on language control and task-switching, our analysis revealed notable similarities across all identified neural areas. Liu and Cao13 also identified the left STG (task switching), right STG (language switching), left culmen (language switching), SMG (task switching, IPL) and MFG/cingulate gyrus (language switching), matching our results. Only the precentral gyrus (LH)13,44 was reported in the opposing hemisphere. This could be a result of thresholding differences (although ours was similar to Liu and Cao13), or differences in anatomical localization or labelling. Another area of difference is in the bilinguals that were included. While we aimed to focus on simultaneous bilinguals, for methodological reasons, we could only use a mixed group of bilinguals. However, 4/6 of our included studies would qualify as “early bilinguals” based on the definition used by Cargnelutti et al.44
Whereas no subthreshold clusters emerged for the Bilingual > Monolingual analysis set, we did find one cluster in the left IFG for our Bilingual ≠ Monolingual analysis. However, the comparison of multidirectional contrasts poses challenges in interpretation. The signature for the left IFG seems to signal importance for this area in language processing and cognition, however, more work is needed to understand differences. As no other ALE analyses has compared language and cognition across bilinguals and monolinguals, we contrast our analysis with ALE comparisons of various bilingual cohorts. Cargnelutti and colleagues’44 meta-analysis of adult bilingual brain representation found no surviving activation clusters between their groups of early and late bilinguals use of L1. However, one cluster in the left IFG was displayed in their L2 conjunction analyses of proficient early and late bilinguals suggesting its importance for bilingual representation.44 To fine-tune our interpretations and identify the functional network where the IFG cluster belongs, we conducted an exploratory, unplanned extension consisting in mapping cluster coordinates onto established probabilistic atlases for the Language, Theory of Mind (ToM) and Multiple Demand (MD) networks (see Supplementary Material S4 for detailed results and methodology).78 For the left IFG cluster identified in the bidirectional analysis, peak and median voxelwise atlas probabilities were highest for the Language network (.383/.274, respectively), with much lower correspondence to the ToM and MD networks. This approach converges with our narrative findings, positioning greater left hemisphere Language network dominance in bilingual language processing and supports the particular relevance of the left IFG to language-mediated cognition in development.
Limitations and future directions
A key limitation of this review includes the inability to conduct a coordinate-based meta-analysis of the fNIRS studies. While fNIRS has become a highly popular method with pediatric populations, given its applicability, ease of use with younger populations and low cost, the studies included here present with substantial methodological heterogeneity. The fNIRS literature currently lacks the standardization seen in other neuroimaging fields, posing several challenges for meta-analysis.
One area of inconsistency lies in the acquisition parameters which varied widely across studies. Key factors include the number of sources and detectors, optode array configurations, and optical properties such as wavelengths used (e.g., 690/830 nm vs. 760/850 nm). Variations in source-detector distances (e.g., 2.5–3 cm) and sampling rates (e.g., 10–50 Hz) also influence signal quality and depth sensitivity. These parameters, often limited by the fNIRS system, can affect spatial resolution and inter-study comparability. Most relevant in terms of the goal of the current study, spatial localization methods also lacked consistency. Optode placements are inconsistent across studies, often based only on approximate 10-20 EEG system points or custom cap designs, making it difficult to precisely align regions of interest (ROIs) across studies. In addition, the absence of consistently reported MNI or Talairach coordinates represents a critical barrier to the coordinate-based meta-analytic approach, as these standardized coordinates are essential for spatially aligning activation patterns. Attempts to infer or reconstruct channel locations from scalp-based placements can introduce considerable uncertainty and error due to individual anatomical variability and the diffuse nature of light propagation, which undermines the spatial precision required for coordinate-based meta-analytic approaches.
Analytical variability further impeded synthesis. No single, generally recommended data processing pipeline exists in fNIRS research; studies employed diverse methods for filtering, motion artifact correction, physiological noise removal, and hemodynamic response function modeling. These analytic decisions can profoundly shape the resulting activation patterns and limit comparability across studies. Inconsistent reporting standards further complicated synthesis, with essential methodological details and statistical outcomes (e.g., precise effect sizes or their variances), critical for meta-analytic calculations, were frequently absent or unclearly presented.
While previous fNIRS meta-analyses have been conducted, they typically included studies with greater methodological consistency or accounted for variability in by severely restricting their scope to a highly circumscribed area of interest - approaches that were incompatible with the broader objectives and the observed variability within our dataset. Given these challenges, we opted for a narrative synthesis to capture the main findings in a tabular format. Future research would benefit from greater methodological standardization and transparent reporting in fNIRS studies (e.g., see emerging guidelines and resources for transparent reporting and open science practices for fNIRS projects79–83), which would allow for stronger cross-study comparisons to facilitate more robust meta-analytic approaches.
The prevalence of fNIRS may in part explain the lower number of studies using alternative neuroimaging techniques such as fMRI – a limitation noted in prior reviews.39 Given this gap in the literature, we opted to perform an exploratory fMRI meta-analysis to better understand this emerging data. However, this meta-analysis was constrained by the inclusion of only six studies, and the lack of overlap in task paradigms. This lack of task consistency introduces considerable interpretive challenges, as the cognitive and linguistic processes engaged by each study are likely to differ, reducing the comparability of activation patterns across datasets.
Another unaccounted source of heterogeneity is individual variability in functional brain network boundaries. Most studies rely on group-level analyses with spatial normalization, which, while standard, can mask meaningful anatomical and functional differences, especially in pediatric samples with variable developmental trajectories. As a result, it is difficult to determine whether similar reported activations reflect truly equivalent underlying neural processes, or whether they arise from coarse averaging across functionally distinct regions. Probabilistic mapping approaches78,84 have been proposed to address these challenges by providing spatially precise network boundaries. However, as these atlases predominantly derive from adult cohorts, they may not accurately capture the dynamic developmental changes in pediatric brains. As the approach relies on existing datasets that vary in acquisition parameters and population characteristics, it can also potentially introduce biases and limit generalizability for underrepresented groups. Nonetheless, probabilistic mapping remains a promising approach, particularly as larger, age-appropriate, harmonized datasets become available.
With a larger evidence base, future research could also focus on untangling finer group differences observed in sub-lexical and lexical and supra-lexical language, as well as cognition, beyond what we are able to do in our current review. Diverse tasks within these categorizations can be expected to elicit distinct neural responses across varying brain regions and functional networks. This lack of overlap in task paradigms within and across groups makes it challenging to attribute observed neural patterns (such as lateralization differences) solely to bilingual type (simultaneous vs. sequential) rather than to the differing cognitive demands of the tasks themselves. This limitation is consistent for the qualitative synthesis of the studies in this review. However, due to the limited scope of the literature, it is not currently feasible to pursue this, especially within tasks that encompass both language and cognitive processing.
In this review, we interpreted potential differences in activation levels across language groups. In the broader neuroscientific context of neural processing, the interpretation of difference in degree of activity remains ambiguous. Whether such activity differences signal enhanced automaticity or efficiency in bilinguals (e.g., through less activity in a region) or greater cognitive effort compared to monolinguals (e.g., through more activity) is not clear.47,85,86 While various theories have been proposed to contextualize findings in young brains (including Johnson’s91 interactive specialization), however, these do not provide direct insights into interpretations of activation levels.
Our critical appraisal of the literature found that SES is not always measured or matched across language groups, which poses a potential issue for bilingual cognitive neuroscience research as SES is a known moderator of task performance.87–89 Only about half of the papers we reviewed reported and controlled for SES. Another challenge of bilingual research is that bilingualism is multidimensional and is measured in different ways depending on the conventions of the field, laboratory, and study.90 This poses a problem when there is not enough information to characterize the participant group in a meaningful way to conduct group comparisons across studies, as remarked in previous reviews.49 Regarding bilingualism factors, both age of acquisition and language proficiency were not consistently and transparently reported. In many cases, when one of these factors was reported, the other was omitted. Age of acquisition was more commonly available or could be inferred from the data, which is why we focused more on this aspect in our review, as it provided more reliable information than proficiency for the studies included. These inconsistencies in reporting SES and bilingual language background motivated our adaptation of the JBI appraisal criteria to account for their inclusion. While various studies lacked information on one or the other, only one study was excluded due to missing information on both SES and bilingual language background. Moving forward, future research should adopt more standardized and transparent reporting practices for bilingual language background and SES to improve cross-study comparisons and strengthen the reliability of bilingualism research.
Conclusion
Taken together, the narrative synthesis of fNIRS and exploratory meta-analysis of fMRI studies revealed similarities between developmental and adult bilingual neural architecture, highlighting a widespread bilateral network that include cortical and subcortical-cerebellar structures. The comparison between monolingual and bilingual children revealed distinct patterns in language and cognition-related brain activation. Simultaneous bilinguals displayed greater frontal and temporal activity during sub-lexical and lexical task processing compared to monolinguals, while sequential/mixed bilinguals exhibited greater activity in left frontal and temporal regions during cognitive tasks compared to monolinguals. Interestingly, sequential/mixed bilinguals showed a closer alignment with monolinguals in left frontal regions during cognitive tasks, indicating potential effects of early vs. later language exposure on functional brain activation. However, given the limited number of studies and variability in participant characteristics and task designs, further research is needed to confirm and expand upon the present preliminary findings. Critically, the field would benefit from the adoption of standardized methodological and reporting guidelines, particularly for fNIRS research, including detailed documentation of acquisition parameters, preprocessing pipelines, and statistical outcomes. Such consistency is essential to improve reproducibility, facilitate meta-analytic integration, and advance understanding of bilingual brain development.
Funding Sources
This research was funded by the Natural Sciences and Engineering Grant (RGPIN-2019-06523) to M.M and the Natural Sciences and Engineering Research Council - Alexander Graham Bell Canada Graduate Scholarship-Master’s to K.I.L.
Conflicts of Interest
The authors have no conflicts of interests to disclose.
{kind=link}