_the_three_r.tiff)
Introduction
The neuroimaging field has made remarkable progress in mapping the brain’s functional architecture. The 2025 Organization for Human Brain Mapping (OHBM) meeting showcased exciting developments: functional gradient approaches, field theories, and continuum models that move beyond discrete network thinking to embrace the brain’s spatial continuity. Yet as our methods evolve, an interpretational gap persists. We are increasingly using eigenmode decompositions — from functional gradients1 to geometric modes2 — but treating eigenmodes as spatial templates with varying weights without a clear understanding of the fundamental principles governing harmonic phenomena. If these patterns truly represent eigenmodes, then to understand their origin and meaning, we must move beyond observational studies or mathematical predictions and look to the physical system itself and the principles that govern eigenmode phenomena in bounded systems in nature — principles well established in acoustics, structural dynamics, vibrational mechanics and fluid dynamics.3,4
The eigenmode revolution
Eigenmodes have become increasingly prominent in neuroimaging research over the past few years. Researchers are discovering that brain activity can be meaningfully described as combinations of eigenmodes. Some approaches extract these eigenmodes directly from observed brain dynamics — principal components or functional gradients of functional magnetic resonance imaging (fMRI) signals. Others derive eigenmodes from structural properties through Laplacian transformations — geometric modes from cortical surfaces, connectome harmonics from structural connectivity.5,6 Others extract eigenmodes at each time point,7,8 or at selected moments when high amplitude events occur.9 While differing in the way the basis of eigenmodes is obtained, these approaches as a whole have successfully predicted functional connectivity patterns, characterised task-evoked responses, and even identified disease signatures.
However, there is a conceptual disconnect in how we discuss these eigenmodes. In the neuroimaging literature, eigenmodes are typically treated as spatial templates with varying weights — mathematical functions that efficiently compress data into a ‘low-rank’ basis. While this mathematical perspective is valid, it overlooks a powerful physical interpretation: eigenmodes in natural systems are fundamentally linked with resonance phenomena.
Every physical system — a drum, a bridge, a bell, and potentially a brain — has its own intrinsic repertoire of resonant patterns called eigenmodes (from the German “eigen” meaning “own” or “inherent”). These aren’t imposed from outside; they’re inherent to the system’s structure, geometry, and physical properties. When you disturb a system (tap a drum, pluck a string, or introduce noise), it doesn’t respond in random ways. Instead, it preferentially activates these natural patterns.
Eigenmodes can be intuitively understood as the system’s repertoire of “dancing movements”—but this is merely an analogy. The eigenmode framework extends far beyond mechanical vibrations. Eigenmodes describe the intrinsic patterns in any system governed by wave equations or diffusion processes: energy states in quantum systems, temperature distributions in thermal diffusion, concentration gradients in chemical systems, and molecular vibrations in proteins. In each case, eigenmodes represent the fundamental ways the system can evolve in space and time.
Crucially, eigenmodes are properties of the system itself as a whole, not of any isolated part within it or particular stimulus. They define what’s possible dynamically, constraining how the system can respond to any perturbation. The specific repertoire of eigenmodes—which patterns exist, at what frequencies, how they’re damped—is determined entirely by the system’s physical properties: its shape, boundaries, material properties, and internal structure.10
Complex dynamics emerge when multiple eigenmodes are excited simultaneously, combining and superposing with each other. This physical interpretation is particularly compelling because of a key mathematical property: eigenvectors are orthogonal to each other. In the context of resonance, this orthogonality has a natural meaning — it describes standing waves with different spatial patterns that can coexist without interacting. Each eigenmode represents a distinct way the system can vibrate, and because they’re orthogonal, they form independent “channels” of oscillation.
While the question remains whether ‘resting-state networks’ reflect eigenmode dynamics or network interactions, bringing concepts from resonance mechanics (typically studied in acoustics and structural dynamics) offers a complementary lens that can bridge the gap between our mathematical methods and their physical interpretation, revealing resonance as a potential self-organizing principle governing the spatiotemporal patterns we observe in neuroimaging.
_spatiotemporal_representation_of.jpg)
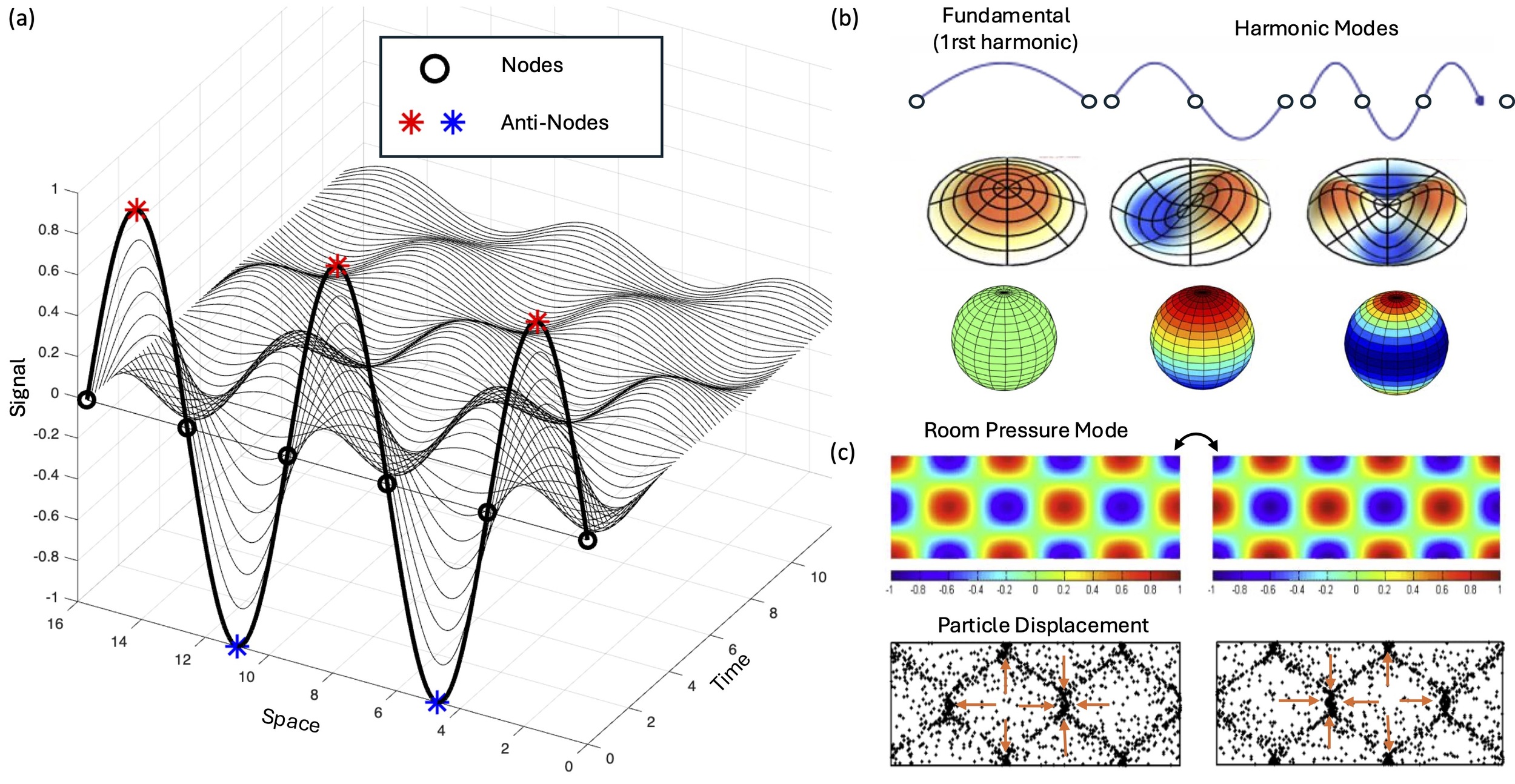
Nodes and antinodes of standing waves
Consider what happens when you pluck a guitar string or strike a drumhead. The resulting standing wave reveals the three defining characteristics of an eigenmode:
-
A characteristic spatial pattern - consisting of nodes (stationary regions of ‘no motion’) and antinodes (regions of maximum displacement that oscillate).
-
A natural frequency - how fast the pattern oscillates when excited; in acoustic systems, we hear a sound when this frequency falls in the range of frequencies we can hear, ranging from 20Hz (low pitch) to 20 000Hz (high pitch).
-
A damping time - how quickly the oscillation fades away over time.
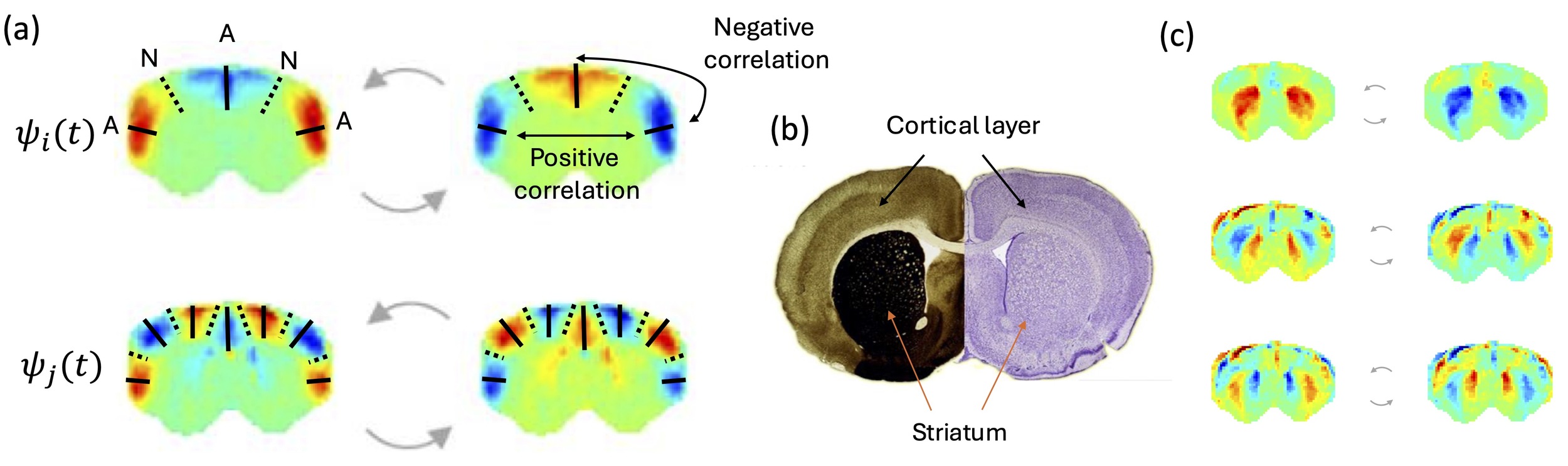
It is crucial to clarify terminology: a “node” in vibrational mechanics is completely different from a “node” in network theory. Wave nodes are stationary regions that can be visualized in vibrating Chladni plates, where sand accumulates precisely because these regions don’t move. Nodes correspond to points in 1D systems like strings, lines in 2D surfaces like membranes, or surfaces in 3D structures like spheres (and brains). Conversely, anti-nodes are the points of maximal variation, where signals oscillate with maximal amplitude. When an eigenmode is excited, the elements in opposite sides of a node vary in antiphase. As such, the phase relationship between antinodes depends on the number of nodes separating them: antinodes separated by an odd number of nodes (1, 3, 5,…) oscillate in anti-phase, while those separated by an even number of nodes (2, 4, …) oscillate in phase.
This spatial organization is not arbitrary — it’s determined by the boundary conditions and physical properties of the system. Critically, this principle offers a geometric explanation for long-range functional connectivity: anatomically distant regions can be strongly correlated if separated by an even number of nodal surfaces, while nearby regions can be anticorrelated if separated by an odd number of nodal surfaces.
If brain dynamics involve standing wave phenomena, then the functional boundaries we observe in neuroimaging may reflect nodes in underlying wave patterns rather than discrete network borders. Regions that appear strongly correlated may be antinodes of the same mode, oscillating together. Anticorrelations may emerge not from inhibitory connections, but from regions lying on opposite sides of a nodal line.
_s.jpg)
From spatial patterns to spatiotemporal dynamics
The shift from viewing brain organization as static functional networks to recognizing it as a repertoire of dynamical modes requires embracing the temporal dimension that conventional fMRI has largely obscured. Recent work using ultrafast fMRI in rodents has revealed that the spatial patterns we identify as “resting-state networks” — now accepted to extend across mammalian brains — actually oscillate with sustained periodicity in the slow frequency range (0.01-0.5 Hz). These are not aperiodic activations following a hemodynamic response function — they show the damped oscillatory behavior characteristic of resonance phenomena.
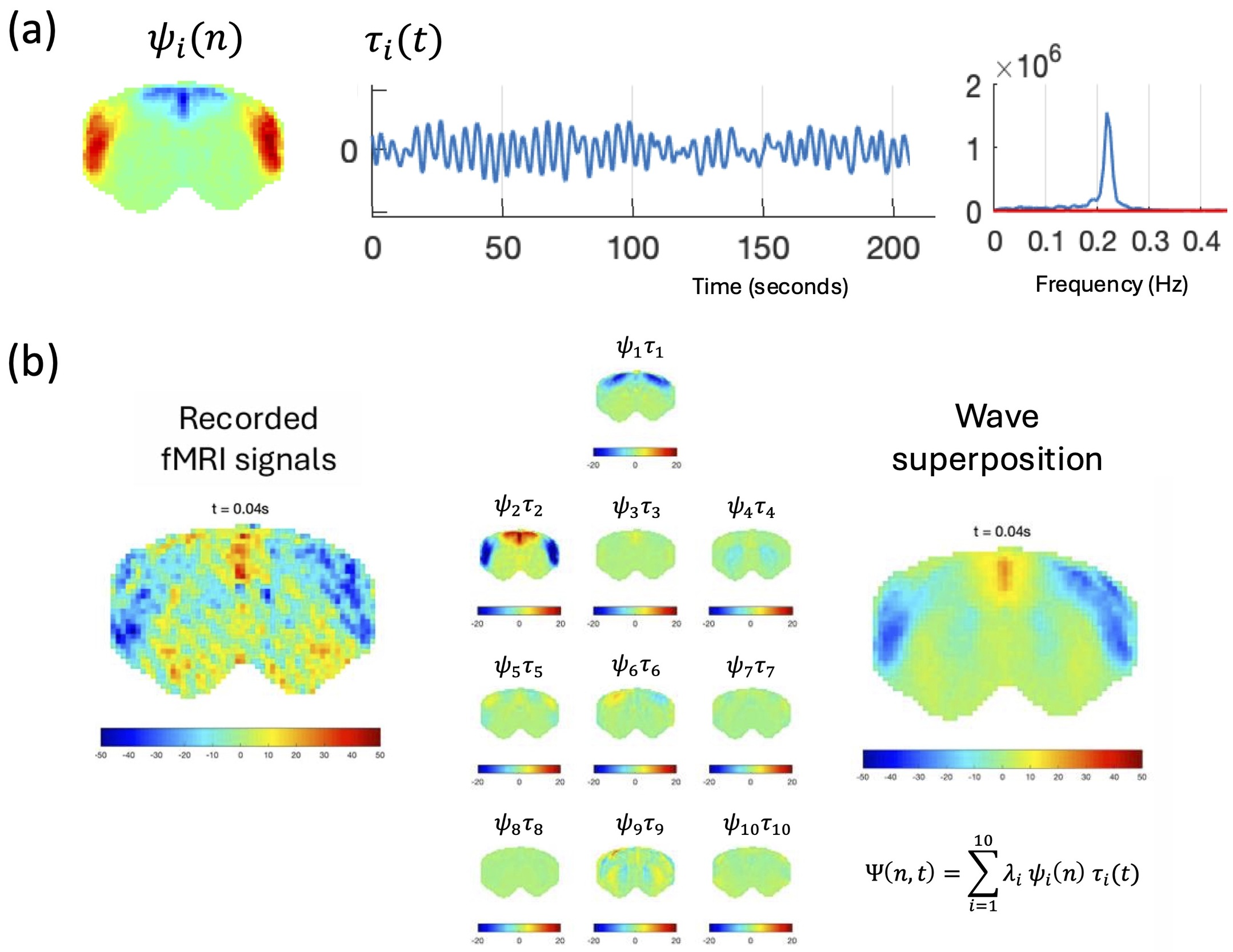
Crucially, what resonates is not a single brain region or node, but an entire spatial pattern spanning the brain simultaneously. A standing wave can be described as a spatial pattern ψ(x) multiplied by a temporal oscillation τ(t) — the whole pattern evolves together as a unit, with each location contributing according to its position relative to the nodes and antinodes. When multiple modes with different frequencies are simultaneously excited, their superposition creates traveling waves through constructive and destructive interference — precisely the propagating patterns increasingly observed in neuroimaging data.11,12
Figure 3 illustrates this decomposition in rat brain data, showing how the recorded fMRI signal at any time point can be reconstructed from the linear superposition of a small number of eigenmodes, each with its own spatial pattern and characteristic oscillation frequency.13
_example_of_.jpg)
A standing wave is an inherently non-local phenomenon: the oscillation belongs to the spatial pattern as a whole, not to any individual location within it. A critical test of this is provided by a spatial shuffle control. When the spatial patterns are randomly permuted across brain locations, the spectral peaks disappear.13 The oscillations are therefore a property of the collective spatial mode, not of individual brain regions.
The observation that resting-state correlations in humans are punctate, occurring in brief windows of a few seconds and less than 5% of the time, is not in tension with the eigenmode resonance view. It is precisely what one would expect from a damped resonant system driven by noise: a resonator does not oscillate continuously, it responds to perturbation, rings briefly (even just a single cycle if critically damped), and then returns to rest. The spatial pattern that emerges during that brief excitation is the eigenmode. This is consistent with the recurrent phase-locking events, termed ‘ghost attractors’ described in Vohryzek et al.8 where brief windows of fMRI phase coherence recur consistently over time and across subjects, drawn repeatedly toward the same spatial configurations without ever fully settling into them. In typical human resting-state fMRI, the system may operate near or below the critical damping threshold, producing the overdamped response visible in Figure 4c: no sustained oscillations, but a spectral preference near the natural frequency. The sustained periodic oscillations visible in rodent ultrafast fMRI may reflect a shift toward underdamped behaviour under medetomidine sedation, which reduces the effective damping coefficient and allows modes to ring for longer. Human resting-state patterns may also reflect the expression of eigenmodes, yet operating in a critically damped regime.
Implications for interpreting brain functional connectivity
Adopting a wave mechanics perspective doesn’t negate network neuroscience, but rather enriches it. Neural connectivity undoubtedly matters, but perhaps not in the way we’ve assumed. Rather than connections directly driving correlations between regions, anatomical structure may define boundary conditions that determine which modes can resonate.
Adopting this framework invites us to reinterpret several foundational assumptions in brain mapping:
-
Anticorrelations need not imply inhibition. Negative BOLD correlations reported for over 20 years14 may simply be antinodes in anti-phase, separated by nodes — a geometric rather than mechanistic relationship.
-
Unlike anatomical boundaries, functional boundaries are not static. Functional boundaries exist only while a mode is excited. They can be recurrent in time and consistent across subjects, without having any detectable anatomical counterpart. The functional boundaries of one mode may correspond to an antinode of another, such that the same anatomical region can simultaneously sit at a boundary in one mode and at a peak of activity in another.
-
Frequency content becomes interpretable. The specific ultra-slow oscillation frequencies detected in fMRI may reflect natural resonances of the brain’s biomechanical system, not just filtered neural rhythms. As suggested by studies showing glymphatic pulsations15 at the frequency range of resting-state activity.
-
State changes reflect physics, not just physiology. Alterations in functional connectivity during different mental states, under anesthesia, or in disease, may involve changes in mechanical properties (stiffness, damping) altering the repertoire of modes that can be excited and how they resonate10,16,17 — offering a physical rather than purely synaptic explanation for why brain dynamics change so dramatically across states.4
-
Eigenmode dynamics cannot be localized. Since eigenmodes are properties of the system as a whole, their origin cannot be assigned to any single region or node. Traditional localizationist approaches — asking which area is “responsible” for a given pattern — are fundamentally mismatched to a framework in which the pattern is the mode itself, spanning the entire system simultaneously.
The role of inhibitory interneurons also takes on a new meaning in this framework. Rather than simply suppressing neuronal activity, local inhibition may act to set nodal lines — by locally reducing excitability at specific locations, inhibitory circuits could shape which spatial patterns are able to resonate and where their boundaries lie. In this view, the classical excitation-inhibition balance operates not just at the level of individual neurons or local circuits, but as a mechanism for tuning the brain’s resonant mode structure at the macroscale.
_the_three_r.tiff)
It is worth noting that physical near-orthogonality arises naturally in linear resonant systems with well-separated eigenfrequencies. In an ideal physical resonator, modes are exactly orthogonal because they represent independent channels of oscillation that do not exchange energy. In a nonlinear, heterogeneous system like the brain, strict orthogonality is an idealisation, and modes are expected to overlap to some degree. Nevertheless, the approximate orthogonality consistently observed in fMRI data is itself informative: it suggests that the brain’s dominant spatiotemporal patterns behave more like the independent modes of a linear resonant physical system than like the entangled outputs of non-linear network interactions.
A critical departure from ideal physical resonators is the pronounced heterogeneity of cortical architecture. The cerebral cortex is not a homogeneous medium: a century of cytoarchitectonic mapping, from Brodmann’s foundational parcellation to modern multi-modal atlases, has established that spatial gradients in myelination, receptor expression, and laminar composition vary systematically across the cortical surface.18,19 In non-uniform physical media, such gradients introduce mode distortions and frequency shifts relative to the predictions of idealised geometric models. Cytoarchitectonic transitions may therefore act as partial reflectors of travelling waves, contributing to the mode boundaries observed empirically and explaining why eigenmode spatial profiles do not map perfectly onto purely geometric predictions. This heterogeneity is not a problem for the framework but an invitation to extend it: the same cytoarchitectonic boundaries that have been mapped for over a century may turn out to encode physical constraints on resonance, linking the oldest tradition in cortical cartography to one of its newest interpretive frameworks.
Resonance minimizes free energy
Every physical perturbation introduces energy into a system. What distinguishes a resonant system from a non-resonant one is not how much energy it receives, but what it does with that energy. In a non-resonant system, energy spreads freely across frequencies and spatial configurations, dissipating rapidly and leaving no lasting coherent trace. A resonant system behaves differently: it selects energy at its natural frequencies and traps it in stable spatiotemporal patterns, its eigenmodes. Rather than dispersing freely, energy is held in coherent oscillating modes whose decay is governed by damping time constants determined by the system’s resonant properties. The Q-factor, familiar from acoustics and electrical engineering, is precisely a measure of how effectively a system ‘traps’ energy in this way, being proportional to the number of sustained cycles before the amplitude decays: a high-Q system sustains oscillations long after the perturbation has passed, because its eigenmodes offer configurations of minimal dissipation.
This physical picture maps directly onto a longstanding theoretical framework in neuroscience. The Free Energy Principle (FEP) proposes that biological systems self-organise to minimise free energy, maintaining structural and functional stability by managing entropy and minimising surprise in their internal states.20 While the FEP is most often cast in computational or statistical terms, its thermodynamic roots are equally fundamental: minimising free energy means directing the system toward configurations where energy is not freely available to do disordered work. And eigenmode resonance could precisely be the physical mechanism by which this is achieved. When brain dynamics conform to the intrinsic eigenmodes of the system, energy is trapped rather than freely dissipated: the system occupies configurations of minimal entropy and maximal coherence.
This connection is supported by recent work showing that Metastable Oscillatory Modes, collective rhythms emerging from transient synchronisation across brain regions,21 are directly associated with reductions in the entropy of ongoing brain dynamics, providing an empirical bridge between brain rhythms and the entropy management central to the FEP.22 The present framework offers the physical grounding for this observation: eigenmodes are the configurations in which the brain most efficiently traps energy, and metastable excursions into these modes are therefore signatures of the system minimising its free energy through resonance. Crucially, this does not replace the synaptic or computational mechanisms described by predictive coding and active inference, but instead describes a possible physical mechanism through which they may operate. Given that brain anatomy and connectivity have boundary conditions favouring stable eigenmodes, the brain might have, through evolution and development,23 built resonance into its physical structure as a strategy for minimising free energy.
The eigenmode framework is primarily demonstrated at the macroscale level of whole-brain dynamics but could be a missing organizing principle operating across scales. While much has been discussed in terms of scale-freeness in the brain, eigenmodes are scale specific, since they encompass the entire collective system. Yet, systems can occur at any scale and resonance, being a universal property, occurring in molecules and atoms, should also occur at the larger and slower scales in cells, ganglia and other subcortical structures, ventricles, and even within cortical layers. Eigenmode analysis at finer scales is an open and promising avenue, extending beyond brain dynamics to all levels of physiological processes involving collective systems distributed in space and evolving in time and technically conceivable using available technologies to record multiple spatial points of a system over time, such as ‘video taping’ at the right spatial and temporal scales (e.g., using a camera for optical imaging, MRI, or microelectrode arrays among others). The key point is that the dynamics do not come from a single point source (so any attempt to localize it will dramatically fail) and is not generated by local mechanisms, so it needs to be analyzed using these frameworks.
Waves shaping connections
What physical medium sustains these eigenmodes remains an open and important question. One possibility is that long-range synaptic connectivity has itself been shaped by resonance efficiency: given that intrinsic functional networks closely approximate geometric modes,6 anatomical wiring may have co-evolved with the geometry of resonant configurations. A complementary possibility is that the relevant medium extends beyond synaptic connections to electromechanical continua (brain parenchyma, glial networks, or even interstitial fluid) whose propagation properties may sustain the macroscopic brain waves detected in fMRI.24,25 The nodal architecture visible in Figure 2, which respects cytoarchitectonic boundaries rather than purely synaptic ones, is consistent with this interpretation. These hypotheses are not mutually exclusive, and their relative contributions represent a key direction for future investigation.
The relationship between eigenmode resonance and synaptic connectivity may itself be circular rather than unidirectional. The classical assumption is that anatomical connectivity drives correlated activity: regions that are wired together tend to fire together. In the eigenmode view, the causal arrow may also run in the other direction. If large-scale resonant modes produce sustained, spatially organised coactivation patterns, then Hebbian plasticity operating on those recurrent patterns would progressively strengthen connections between regions that consistently coactivate as antinodes of the same mode. Over developmental timescales, the connectome may thus be partly shaped by the resonant geometry of the brain rather than the other way around. A further possibility is that eigenmode dynamics modulate local neuronal excitability rather than driving synaptic firing directly, through spatiotemporal gradients of pressure, ionic concentration, or membrane potential that determine which neurons are near threshold at any given moment. In this view, the eigenmode acts less like a command signal and more like a tide, setting the conditions under which local synaptic dynamics unfold. Whether this constitutes a causal mechanism, a permissive scaffold, or an epiphenomenon remains to be determined, and we present it as one of the most compelling open questions raised by this framework.
It is important to emphasise that the eigenmode resonance framework is proposed as a complementary physical interpretation, not a replacement for mechanistic accounts rooted in synaptic connectivity. Neural firing, synaptic transmission, and local circuit dynamics remain essential components of brain function. The wave mechanics perspective asks a different question: given all of this synaptic machinery, what physical constraints does the brain as a bounded structure impose on the patterns it can express? Resonance and network interaction are not mutually exclusive; they operate at different levels of description, and both are likely necessary for a complete account of brain dynamics.
This eigenmode resonance perspective also carries direct implications for brain stimulation. If eigenmodes are configurations of minimal dissipation, then stimulation tuned to match the brain’s intrinsic resonant frequencies and spatial patterns should produce maximal and sustained effects with minimal energetic input. In other words, rather than resisting the perturbation, the brain would amplify and sustain it through resonance. This offers a physically principled hypothesis for testing stimulation protocols where coherent spatiotemporal patterns are enhanced when stimulated at resonant frequencies in ways that off-resonance stimulation cannot achieve.26
Cognitive and behavioural differences
The eigenmode framework is also opening new avenues for characterising cognitive and behavioural differences within and across individuals. Growing evidence is revealing that the dominance and spatial profiles of eigenmodes vary systematically across conditions, capturing aspects of brain organisation not accessible through traditional localizationist or connectivist methods. Tools such as LEiDA (Leading Eigenvector Dynamics Analysis) already operationalise eigenmode dynamics at the population level, enabling comparison of mode profiles across cognitive traits7,27 and clinical status.28,29 Shifts in the mode repertoire may serve as neuroimaging phenotypes in large cohort studies, with potential relevance to psychiatric syndromes and neurological conditions, including schizophrenia, and depression.30,31
Moving forward
While the present framework is demonstrated at the macroscale of whole-brain dynamics, resonance is a universal physical property and eigenmodes should arise at any scale where a collective system is distributed in space and evolves in time — from cortical layers and subcortical nuclei to individual cells and molecular assemblies.
The neuroimaging community has developed sophisticated methods to identify and characterise spatial modes in brain activity. Now we must bring the full physics of wave phenomena into our interpretations. This means reconsidering that the slow spontaneous oscillations driving spatial patterns of functional connectivity may reflect the brief excitation of an eigenmode — not a region switching on, but an entire pattern resonating. It means treating functional boundaries as potential nodal lines, and apparent traveling waves as the natural consequence of mode superposition.
Crucially, integrating dynamical parameters into existing network-based pipelines is practically straightforward, given that eigendecompositions are already the substrate of most decomposition methods, making translation between frameworks computationally accessible. Rather than replacing network approaches, eigenmode analysis supplies a physical interpretation of how the patterns that network methods describe actually arise. It means expanding our models to include biomechanical properties — stiffness, damping, geometry — alongside synaptic connectivity as determinants of brain dynamics.
The eigenmode resonance framework generates testable, falsifiable predictions that distinguish it from a purely mathematical description. First, if resting-state patterns reflect genuine resonant modes, then stimulation tuned to match both the spatial profile and frequency of a target eigenmode should produce effects that are qualitatively different from off-resonance stimulation. Concretely, transcranial alternating current stimulation or focused ultrasound applied at an antinode, phase-matched to the ongoing oscillation, should selectively enhance that mode’s amplitude, while stimulation at a nodal line should have negligible effect. Comparing tACS and focused ultrasound at matched frequencies would further help disambiguate whether the relevant medium is primarily electrical or mechanical. Second, if eigenmodes are shaped by brain geometry, then systematic changes in morphology across development and across species should produce predictable shifts in the spatial profile of dominant modes. Cross-species comparative neuroimaging, combined with geometric modelling, provides a direct test of this structural prediction. Third, multimodal recordings combining ultrafast fMRI with EEG or MEG would allow direct comparison of electrophysiological and haemodynamic signatures of the same putative modes, helping to determine whether the ultraslow oscillations detected in fMRI reflect a genuine resonant property of the brain or an artefact of neurovascular filtering.
Convergent evidence from wide-field calcium imaging supports the case that eigenmode-like spatial organisation is not an artefact of haemodynamic filtering. Spontaneous patterns of intracellular calcium concentration share the same macroscale spatial structure as resting-state fMRI modes, following geometric rules rather than the discrete boundaries of anatomical connectivity.32–34 That the same geometric organisation appears independently across imaging modalities suggests that both signals are capturing a common phenomon viewed through different physical lenses.
Perhaps the most direct test of whether eigenmode-like spatiotemporal patterns can arise from purely mechanical resonance would not require a biological preparation at all. A physical phantom reproducing the key biomechanical features of the brain,35 such as a deformable medium enclosed within a rigid fluid-filled cavity, could be used to test whether fMRI detects eigenmode-like spatiotemporal patterns driven by mechanical pulsations alone. If such patterns emerge, this would establish that geometry and boundary conditions are sufficient to generate them, providing a proof of concept for the resonance hypothesis and a critical null hypothesis against which biological data can be compared. Such an experiment would also allow direct manipulation of the medium’s mechanical properties, such as stiffness, viscosity and surface tension, to test how each shapes the eigenmode repertoire, something hard to study in a living system.3
Our field is already using the mathematics of eigenmodes. It is time we embrace the physics that comes with them. The patterns we observe in fMRI may be telling us something profound about the brain as a physical system: not just a neural network, but a resonant structure whose dynamics emerge from the interplay of geometry, mechanics, and biology.
The standing wave framework doesn’t answer all questions about brain function — and it may ultimately turn out to be wrong. But science has never advanced by converging prematurely on a single interpretation. In 2026, a growing number of researchers are choosing to approach the brain not only as a network of communicating regions, but as a resonant physical structure whose dynamics emerge from the interplay of geometry, mechanics, and biology. We may be mistaken. But by allowing ourselves the freedom to explore different paths, to ask: 'what the brain sounds like, not just how it is wired’, we open new avenues for understanding how the physical brain gives rise to the dynamics we measure, and ultimately, to the mind itself.
Funding sources
JC is supported by LARSyS through funding by the Portuguese Foundation for Science and Technology (FCT) (doi: 10.54499/LA/P/0083/2020, 10.54499/UIDP/50009/2020 and 10.54499/UIDB/50009/2020)l and Arqus Research Alliance.
Conflicts of interest
The authors declare no conflicts of interest.